冬春季狍取食营养策略及影响因素研究
2023-02-22姜广顺
齐 琦,包 衡,2,姜广顺,2,3
(1.东北林业大学野生动物与自然保护地学院,哈尔滨,150040;2.国家林业和草原局猫科动物研究中心,东北林业大学,哈尔滨,150040;3.东北亚生物多样性研究中心,东北林业大学,哈尔滨,150040)
有蹄类动物会对觅食做出决策,权衡食物质量和数量,食物选择受到植物的形态、数量、分布和次生代谢产物的影响[1-2]。营养生态学关注环境与动物摄取食物之间的营养联系以及由此产生的生理状态[3]。满足自身的营养需求是有蹄类动物行为的主要驱动因素之一,尤在经历繁殖期以及食物资源匮乏和恶劣天气的时期,它们的营养状态决定了生存状态,也决定了其是否能主动取食及取食数量。动物能够适应环境并在野外生存的最基础的条件是取食,只有摄入了足够的食物和均衡的营养,才能够维持自身的最佳身体状况,从而提高其繁育能力和生存能力[4]。目前,营养生态学领域涉及最优觅食理论、经典昆虫营养生态学理论、营养几何框架和生态化学计量学4个理论框架[5-8]。
野生动物的取食和营养策略是由多种因素共同驱动的,包括资源限制和摄取目标位置变化的调节反应[9]。由于体重、代谢适应、生活史和生理状态的变化,不同的个体在选择食物和营养策略时可能会优先考虑不同的食物成分[10-11]。此外,动物摄取营养还受到营养需求的季节性变化驱动,有研究证明动物通过增加夏季和秋季非蛋白能量的摄入量来储备脂肪,以弥补寒冷的冬季能量摄入的限制[12]。关于有蹄类动物的营养策略现有6 种理论假说,即能量摄取最大化假说、蛋白摄取最大化假说、摄食纤维限制假说、矿物质元素限制假说、植物次生代谢物限制假说和营养平衡假说[13]。
狍(Capreolus pygargus)属于哺乳动物鲸偶蹄目(Cetartiodactyla)[14]鹿科(Cervidae)狍属(Capreolus),主要以灌木的枝芽、树叶和各种青草为食,是东北地区常见的偶蹄类野生动物之一,也是大型濒危食肉动物东北虎(Panthera tigris altaica)和东北豹(P.pardus orientalis)的主要猎物之一。狍会随季节的变化调整生理状况和取食策略以应对不同营养环境的挑战[15]。本研究从营养生态学方面探究冬季(11 月前后)和春季(5月前后)珲春地区狍取食营养策略及影响因素。主要拟解决的科学问题:(1)冬春季狍食物组成和营养摄取是否存在差异?(2)食物可获得性是否影响冬春季狍食物组成,以及可食植物营养含量是否会影响冬春季狍取食该种植物的比例?(3)环境中可食植物宏营养和微营养总量是否影响冬春季狍营养摄取?
1 研究区概况
在吉林省珲春地区开展与狍相关的野外调查和样本采集工作,珲春位于吉林省延边朝鲜族自治州东部的中、俄、朝3 国交界地带。研究地区属于近海中温带季风气候区,气温受到海洋性气候的影响变化比较明显,年平均气温为5.65 ℃,极端最高气温为36.3 ℃,极端最低气温为-32.5 ℃,封冻期在11月初,一般4 月上旬开始解冻。珲春是我国野生东北虎和东北豹分布数量与密度最高的地区,其独特的地理位置和区位优势,使这里成为中、俄、朝3 个国家虎豹种群迁移和扩散的生态廊道,是我国野生东北虎种群向内陆区域扩展的希望所在。
2 研究方法
2.1 数据采集
2020 年11 月—2021 年5 月,在珲春东北虎国家级自然保护区开展植物和狍粪便样本的采集工作。采用系统抽样的方法,把保护区分成100个2 km×2 km的网格,剔除14 个位于村庄和道路上的网格,将余下的86 个网格设置30 m×30 m 的植物样方,记录样方编号、中心点坐标、隐蔽级及乔木(树高>2 m,胸径≥5 cm)种类、数量和平均胸径等信息,在每个植物样方的四角和中心位置共设置5个2 m×2 m的小样方,记录每个小样方内狍可取食高度的灌木(高度0.5~2.0 m,胸径<5 cm)种类、数量、平均枝条数及草本(高度<0.5 m,幼苗与草本共同记录)种类和盖度等信息,并采集狍主要取食的植物样本。返回驻地后及时将植物样本称重,装入档案袋中,记录植物名称、采集地点、植物重量、档案袋重量和采集时间等信息。在采集植物过程中采集遇到的狍新鲜粪便,装入封口袋中,记录采集时间、地点和物种等信息,尽快放入冰箱中,-20 ℃冷冻保存,以便后续分析。
2.2 食性分析和取食植物营养含量测定
采用粪便显微分析法分析狍的食物组成[16]。采用重量法测定粗水分质量分数[粗水分质量分数=(鲜重-干重)×100%];采用凯氏定氮法(FOSS,Kjeltec 8400 全自动凯氏定氮仪)测定粗蛋白质量分数(粗蛋白质量分数=含氮量×6.25);采用纤维分析法测定酸性洗涤纤维(acid detergent fiber,ADF)、中性洗涤纤维(neutral detergent fiber,NDF)、酸性洗涤木质素(acid detergent lignin,ADL)、纤维素和半纤维素含量(ANKOM 2000i 型全自动纤维分析仪);采用索氏抽提法测定粗脂肪含量(全自动固液萃取仪);采用灰化法测定植物样品的粗灰分含量。采用电感耦合等离子体发射光谱(ICPS-7500)ICP-AES 法测定微营养含量。
2.3 数据处理
2.3.1 食物组成和可获得性分析
为了比较冬春季环境中狍食物可获得性对其食物组成的影响,计算狍食物可获得性。分别统计野外每个植物样方中狍每种可食植物枝条数并计算其在每个样方中占所有植物枝条的比例,环境中狍每种可食植物比例为所有样方中每种植物比例总和,即为食物可获得性。
2.3.2 可食植物物种多样性指数和生态位宽度分析
为了比较冬春季狍取食食物组成比例的差异,分别计算狍可食植物物种的丰富度指数(D)、香农-维纳指数(H')、物种均匀度指数(J')和生态位宽度(B),计算公式如下:
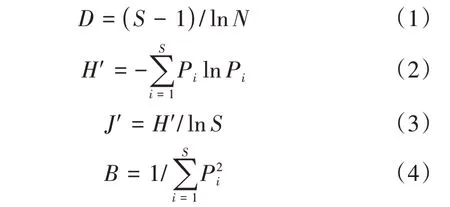
式中:S表示可食植物物种的总数目;N表示所有可食物种的总体个数;Pi表示第i个物种的个体数。
2.3.3 宏营养和微营养的摄取及供应分析
宏营养包括蛋白质、脂肪、总结构性碳水化合物(total structural carbohydrates,TSC)、总非结构性碳水化合物(total non-structural carbohydrates,TNC)和总能(gross energy,GE),微营养主要包括钠(Na)、磷(P)和钙(Ca)。TSC 包括纤维素、半纤维素和木质素,由于木质素在动物体内不可被消化,因此TSC 质量分数=纤维素质量分数+半纤维素质量分数;TNC质量分数=100%-(粗蛋白质量分数+粗脂肪质量 分数+NDF 质量分数+粗灰分质量分数)。每100 g 植物中所含的总能=23.64 kJ/g×蛋白质质量分数+39.54 kJ/g×粗脂肪质量分数+17.15 kJ/g×(TSC 质量分数+TNC质量分数)[17]。
狍从食物中摄取的营养为植物中营养含量乘以该种植物在狍食物组成中所占比例。营养供应分为可食植物营养供应和可食植物归一化后可供应营养,其中,可食植物营养供应为植物中营养含量乘以环境中该种植物所占比例;可食植物归一化后可供应营养为可食植物营养供应乘以该种植物在狍食物组成中所占比例。应用单样本秩和检验(U检验)比较冬春季狍营养摄取和可食植物营养供应之间的差异。
3 结果
3.1 狍的食物组成和可获得性
在珲春地区共收集到狍粪便38 份,其中冬季26份,春季12 份;共收集到植物样本953 份,其中冬季542 份,春季411 份。经统计显示:冬季狍食物组成比例为山核桃属(Caryaspp.,14.02%)、栎属(Quercusspp.,11.18%)、桦木属(Betulaspp.,10.41%)、槭 属(Acerspp.,10.34%)、椴树属(Tiliaspp.,9.15%)、榆属(Ulmusspp.,7.92%)、松属(Pinusspp.,7.10%)、杨属(Populusspp.,4.40%)、桤木属(Alnusspp.,3.67%)、溲疏属(Deutziaspp.,3.44%)、榛属(Corylusspp.,3.02%)、忍冬属(Loniceraspp.,2.34%)、柳属(Salixspp.,1.73%)、卫矛属(Euonymusspp.,1.17%)、绣线菊属(Spiraeaspp.,1.08%)、稠李属(Padusspp.,0.74%)和丁香属(Syringaspp.,0.25%);春季狍食物组成比例为松属(27.40%)、柳属(18.43%)、槭属(17.38%)、桦木属(4.04%)、椴树属(3.69%)、榛属(2.93%)、苹果属(Malusspp.,2.93%)、杨 属(2.67%)、薹草属(Carexspp.,2.55%)、鳞毛蕨属(Dryopterisspp.,2.45%)、忍冬属(2.26%)、绣线菊属(2.36%)、梨属(Pyrusspp.,2.01%)、荚蒾属(Viburnumspp.,1.85%)、杜鹃花属(Rhododendronspp.,1.11%)、卫矛属(0.76%)、丁香属(0.54%)、山楂属(Crataegusspp.,0.67%)、鼠李属(Rhamnusspp.,0.48%)、稠李属(0.48%)、胡枝子属(Lespedezaspp.,0.32%)、茶藨子属(Ribesspp.,0.22%)、云杉属(Piceaspp.,0.22%)、榆 属(0.19%)、栎属(0.16%)、五味子属(Schisandraspp.,0.10%)、桤木属(0.16%)、小檗属(Berberisspp.,0.16%)、苔藓(Bryophyta,0.10%)、合欢属(Albiziaspp.,0.10%)、五加属(Eleutherococcusspp.,0.10%)和珍珠梅属(Sorbariaspp.,0.03%)(图1A)。
冬春季环境中狍食物可获得性计算结果表明,在冬季,榛属、槭属、忍冬属、柳属、栎属、松属、桦木属和丁香属的可获得性较高;在春季,薹草属、槭属、榛属、忍冬属、柳属、胡枝子属、鳞毛蕨属和松属的可获得性较高(图1B)。
通过对比狍食物组成和可获得性结果可知,冬季可获得性较高的槭属和栎属在其食物组成中占比较大,但榛属、忍冬属和柳属在其食物组成中占比并不高,并且食物组成中占比较高的山核桃属、桦木属、椴树属和榆属可获得性较低;春季可获得性较高的槭属在其食物组成中占比较大,但薹草属、榛属和忍冬属在其食物组成中占比并不高,并且食物组成中占比较高的松属和柳属可获得性较低(图1)。
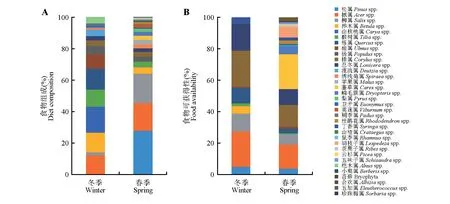
图1 冬春季狍食物组成与食物可获得性差异Fig.1 Comparison of diet composition and food availability of Capreolus pygargus in winter and spring
3.2 冬春季狍可食植物物种多样性指数和生态位宽度
表1 显示,冬季狍取食植物的物种丰富度指数、香农-维纳指数、均匀度指数和生态位宽度均低于春季,说明相比于冬季,春季狍的食物数量更多,食物选择更为泛化。

表1 冬春季狍食物物种多样性指数及生态位宽度Tab.1 Food species diversity index and niche breadth of Capreolus pygargus in winter and spring
3.3 冬春季狍宏营养摄取和取食植物宏营养供应差异
3.3.1 每种食物宏营养供应差异
通过冬春季环境中狍每种食物宏营养(蛋白质、脂肪、TSC、TNC和GE)供应比例的计算结果可知,冬季环境中食物宏营养供应主要以槭属、栎属、榛属、桦木属、忍冬属和松属为主,其中槭属、栎属、桦木属和松属在狍食物组成中所占比例较大,另外脂肪供应较多的椴树属在狍食物组成中占比较大,GE供应较少的忍冬属在食物组成中占比较小,可见冬季狍主要选择脂肪和GE供应较多的食物;春季环境中食物宏营养供应主要以槭属、薹草属、柳属和松属为主,其中槭属、柳属和松属在狍食物组成中所占比例较大,而薹草属在狍食物组成中所占比例较小(图2)。
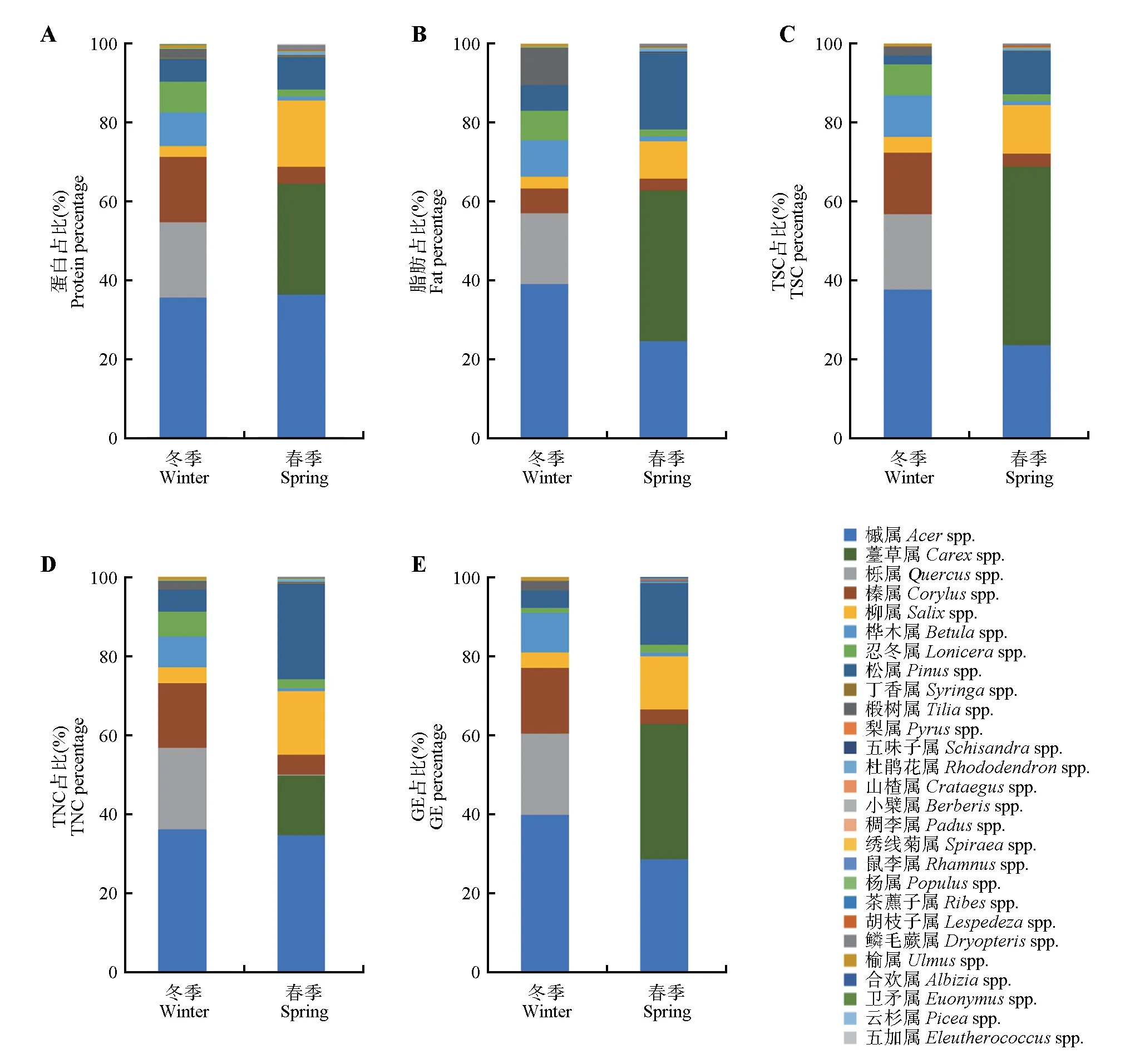
图2 冬春季环境中狍每种食物宏营养供应差异Fig.2 Comparison of macro-nutrition supply for Capreolus pygargus in winter and spring
3.3.2 宏营养摄取及供应差异
对冬春季狍摄取的宏营养进行差异性分析的结果表明,冬季狍从植物中摄取脂肪、TSC、TNC 和GE含量均显著高于春季(P<0.05),冬春季狍从植物中摄取蛋白质(P=0.08)的含量无显著差异(图3A),表明冬季温度低,狍主要摄取与能量有关的营养。
对冬春季环境中狍取食植物的宏营养供应进行差异性分析的结果表明:冬季环境中狍取食植物脂肪(P=0.11)、TSC(P=0.09)、TNC(P=0.11)和GE(P=0.17)含量高于春季但差异不显著,蛋白质(P=0.45)含量低于春季但差异不显著(图3B)。
对冬春季环境中狍取食植物归一化后可供应宏营养进行差异性分析的结果表明,冬季环境中狍取食植物归一化后可供应脂肪、TSC、TNC 和GE 含量均显著高于春季(P<0.05),蛋白质(P=0.13)含量低于春季但差异不显著(图3C)。通过对比上述结果发现,狍对于宏营养的摄取及环境中宏营养供应在蛋白质、脂肪、TSC、TNC 和GE 含量上的变化一致,说明冬春季环境中狍取食植物宏营养的供应对其摄取量存在影响。
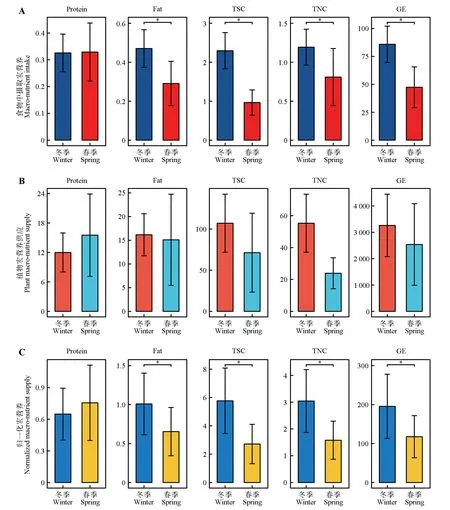
图3 冬春季狍宏营养摄取、取食植物宏营养供应、取食植物归一化后可供应宏营养差异Fig.3 Differences in macro-nutrient intake,feeding plant macro-nutrient supply and feeding plant normalization of Capreolus pygargus in winter and spring
3.4 冬春季狍微营养摄取和取食植物微营养供应差异
3.4.1 每种食物微营养供应差异
通过计算冬春季环境中狍每种食物微营养(Na、P 和Ca)的供应比例发现,冬季环境中食物微营养供应主要以槭属、栎属、榛属、桦木属和忍冬属为主,其中槭属、栎属和桦木属在狍食物组成中所占比例较大;春季环境中食物微营养供应主要以槭属、薹草属、柳属和松属为主,其中槭属、柳属和松属在狍食物组成中所占比例较大(图4)。
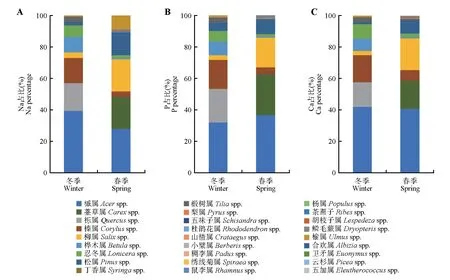
图4 冬春季环境中狍每种食物微营养供应差异Fig.4 Comparison of each food micro-nutrient supply for Capreolus pygargus in winter and spring
3.4.2 微营养摄取及供应差异
对冬春季狍摄取微营养进行差异性分析的结果表明,冬季狍从植物中摄取Na、P 和Ca 含量均显著高于春季(P<0.05)(图5A),说明冬季狍对于Na、P和Ca的需求更高。
对冬春季环境中狍取食植物微营养供应进行差异性分析的结果表明,冬季环境中狍取食植物Na(P=0.32)和P(P=0.49)含量低于春季,Ca(P=0.43)含量高于春季,且差异均不显著(图5B)。
对冬春季环境中狍取食植物归一化后可供应微营养进行差异性分析的结果表明,冬季环境中狍取食植物归一化后可供应Na(P=0.10)、Ca(P=0.07)含量均高于春季,P(P=0.14)含量低于春季,且差异均不显著(图5C)。
通过对比上述结果发现摄入量和供应量在Ca含量上的变化一致,P 含量上的变化相反,而冬春季狍摄入Na含量与取食植物归一化后微营养供应变化一致,与取食植物微营养供应变化相反(图5),说明冬春季环境中狍取食植物P和Ca的供应对其摄取量存在一定的影响。
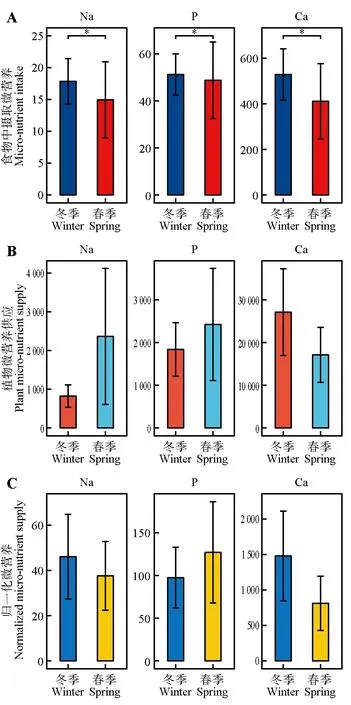
图5 冬春季狍微营养摄取、取食植物微营养供应、取食植物归一化后可供应微营养差异Fig.5 Differences in micro-nutrient intake,feeding plant micro-nutrient supply and feeding plant normalization of Capreolus pygargus in winter and spring
4 讨论
4.1 食物组成影响因素
狍是对食物具有高度选择性的草食动物,可食植物质量和数量是其生存和繁衍的主要影响因子,也是影响其分布和栖息地选择的主要驱动因子[18-20]。狍对于食物的选择会随季节和纬度的变化而变化[21],冬季狍的食物数量和质量均处于一年当中的最低水平[22]。本研究表明,冬春季狍取食植物种类存在差异,冬春季的植物可获得性也存在差异,环境中可获得性高的食物在狍食物组成中所占比例不一定高。驼鹿(Alces alces)营养适应策略的研究说明食物可获得性对决定营养摄入量的平衡不是关键,是由于食物的质量和重量的共同作用使它们有机会在冬季平衡饮食[23]。因此,食物可获得性对于狍食物组成比例并不是主要决定因素,食物可获得性及其各种营养含量共同影响决定了狍冬春季食物组成。
另外,狍取食植物种类的多少受到食物物种丰富度的影响,在不同的季节受影响的程度也不同[24],由于冬季可利用的植物减少,狍种内取食植物的相似度提高[25],冬季狍取食植物的多样性指数均低于春季,说明相比于冬季,春季狍的食物数量更多、食物选择更为泛化。
4.2 营养摄取策略
根据最优觅食理论,获得优质食物是对动物种群动态进行调节的关键性因素之一。在食物和营养资源充足的情况下,动物优先选择取食养分高的植物;在食物和营养资源匮乏的情况下,则权衡植物的营养成分和生物量来进行选择[26],并且最大限度地提高能量的净摄入量[27]。有研究表明,在植物的生长季节,动物会选择一些可利用生物量低但营养含量高的食物[28]。本研究发现,在食物和营养资源缺乏的冬季,狍主要选择取食可获得性且养分含量高的植物(如槭属、栎属)和能量供应量高但可获得性中等的植物(如桦木属、椴树属和松属);在食物和营养资源充足的春季,狍对于可获得性高的食物(如薹草属、忍冬属和榛属)取食较少,而主要选择取食养分含量高的植物(如槭属、柳属和松属)。
植物的营养多样性程度对食草动物的适合度有很大的影响,动物能够有选择地取食植物并管理各种抑制其取食的植物特征[29]。蛋白质在动物的生长发育、代谢和繁殖中起着重要作用,在北方生态系统中,植物的蛋白质含量通常在生长季节达到峰值[30]。Felton 等[13]的研究显示,没有足够的证据表明鹿科动物在夏季对蛋白质的选择比冬季更强烈,本研究中冬春季狍对蛋白质的摄入量无显著差异,符合Felton 等的研究结果。动物主要的能量来源于脂肪和碳水化合物,对于特定的生命阶段,维持日常代谢所需要能量基本上是固定的,但由于活动、繁殖和天气等因素的影响需要摄取额外的能量[31]。高纤维饮食比低纤维饮食在消化过程中产生了更多的热量,因此,动物在较低的温度下会摄取更多的纤维[32]。本研究发现冬季环境中脂肪、TSC、TNC 和GE 的供应量和狍摄取量均高于春季,这说明为了适应冬季寒冷的天气、满足繁殖和体温调节的需求,狍会有选择地摄取更多能量高的植物。与宏营养相比,动物对微营养(Na、P和Ca)的需求量较少,但其对骨骼健康至关重要,在许多代谢过程中是必不可少的[33]。狍为了适应寒冷的天气和维持正常的代谢,冬季对于Na、P和Ca的需求更高。
4.3 取食策略的影响因素
由于冬季和春季气候因子的变化,植物的数量和质量都会发生改变,动物会通过改变取食行为和营养策略来适应不同的气候变化[34]。动物会将营养互补的食物组合在一起,组成具有与目标食物非常相似的常量营养素平衡的食物组合[35]。本研究表明,冬春季植物营养含量发生变化,狍会选择不同的食物组合达到摄入的营养平衡从而满足自身在不同季节的营养需求。在低生物量水平下,动物的能量摄入可能会受到可食植物数量的限制,在中等生物量水平下,其能量摄入可能会受到可消化能量和蛋白质含量的限制[19,36]。因此,在冬季狍可食植物的种类和数量较少,为了满足自身的能量需求会选择取食碳水化合物含量高的植物来增加食物在体内的消化时间以减少食物的摄入量,而春季可食植物的种类和数量增加,其能量摄入则是通过综合考虑蛋白质和可消化能量获得的。由于北方生态系统冬季气温较低且有大量积雪,动物消耗的能量高于春季,为了平衡其热量损失和能量消耗,会增加含能营养素(脂肪和碳水化合物)的摄入量,冬季脂肪和碳水化合物的摄入量增加将与额外的体温调节成本相匹配[34]。微营养对于动物体内的物质代谢及维持生理功能至关重要,冬季狍从植物中摄取更多的Na、P 和Ca,可能与适应寒冷的环境和维持正常代谢及生理功能有关。
致谢:感谢珲春林业局对本研究野外调查工作的支持和帮助。
