四明山野生杜鹃花资源遗传多样性和亲缘关系分析
2023-02-20李东宾徐婧何立平许志斌沈登锋
李东宾 徐婧 何立平 许志斌 沈登锋
(1.浙江省宁波市林场,浙江 宁波 315440;2.宁波市农业科学研究院,浙江 宁波 315040)
杜鹃花属(Rhododendron)是北半球最大的木本植物属,属内植物大多株形古朴优雅、花色丰富艳丽,具有极高的观赏价值,是我国十大传统名花,在世界园艺界也占有重要地位[1]。中国是野生杜鹃花属植物资源最丰富的国家,有720种,其中特有种450种[2]。浙江省杜鹃花资源有17种2变种及1变型(不包括栽培种)[3],浙东四明山区域有自然分布的为映山红(R.simsii)、满山红(R.mariesii)、华顶杜鹃(R.huadingense)、羊踯躅(R.molle)、马银花(R.ovatum)、云锦杜鹃(R.fortunei)6种,隶属于4个亚属[4]。
遗传多样性是生物多样性的本质,是物种适应环境的核心基础和进化的内在动力,也是保护生物学研究重点工作,可为物种保护及利用提供适宜的策略[5]。随着分子标记技术体系日益成熟,利用分子标记技术探究评估种质资源的遗传多样性和亲缘关系已成为一种常用方法[6]。EST-SSR分子标记随转录组测序技术的成熟开发经济、快速、简单,具有信息量大、共显性、通用性高等特点[7],已被广泛运用于龟峰山杜鹃花[8]、西双版纳苦茶[9]、邵武碎铜茶[10]、贵州刺梨[11]、额济纳湿地多枝柽柳[12]等种质资源的遗传多样性研究和亲缘关系分析,并取得了良好的实验结果。
四明山区域地处浙东宁波市西南 部,位 于121°0′31″~121°25′33″E,29°32′13″~29°59′56″N,光照充足,雨量充沛,属于典型的北亚热带气候[13]。区域内地形复杂,山势险峻,沟谷纵横,植物资源丰富,是宁波最重要的生态保护区和水源涵养地。四明山区域分布着国内最大规模的华顶杜鹃野生种群,华顶杜鹃为浙江省特有种,观赏价值高,自然分布区域狭窄,种群数量稀少,并且在系统发育研究中具有重要价值[14],已被列为浙江省珍稀濒危植物、国家二级重点保护野生植物。华顶杜鹃1990年才被发现于天台华顶山,因独特的形态特征其分类归属尚存争议,颜士辉[15]、李美芹[16]等做了华顶杜鹃的分子标记开发及亲缘关系分析,均未能明确解决它的分类归属问题。本文利用EST-SSR标记进行四明山区域内野生杜鹃花资源的遗传多样性和亲缘关系研究,从分子角度分析区域内杜鹃花资源的种群遗传多样性和亲缘关系以及华顶杜鹃的分类归属问题,旨在为该区域内野生杜鹃花的资源保护、种群恢复、种质创新提供科学依据。
1 材料与方法
1.1 材料
本研究对象为在四明山区域有自然分布的映山红、满山红、华顶杜鹃、云锦杜鹃、马银花和羊踯躅6种野生杜鹃花资源。测试样品于2021年9月在四明山国家森林公园内海拔600~800m野生杜鹃花分布比较集中的区块内寻找目标野生杜鹃花资源并取样,每个种采样15个单株,单株距离要大于30m,每株采集3~5片健康幼嫩叶片,迅速置于盛有变色硅胶的密封袋干燥保存。采集完成后带回实验室用液氮处理后存于-70℃超低温冰箱,用于DNA提取。
1.2 DNA提取与质量检测
采用改良CTAB法[17]提取试验样品总DNA,用1.0%的琼脂糖凝胶电泳进行杜鹃花DNA分子量检测,用紫外分光光度计(nano drop 2000)检测提取的DNA的质量和浓度,最终统一将不同样品的DNA浓度稀释至50ng/μL于-20℃放置备用。
1.3 EST-SSR引物的筛选
参照红马银花的12对多态性好且稳定的EST-SSR引物序列[18]进行本试验引物的筛选,每种杜鹃花随机选择3个基因组DNA作为模板对12对引物进行筛选,选取条带扩增清楚,多态性高的引物用来进行试验材料的群体遗传多样性分析。引物由上海生物工程公司合成,筛选出的引物在正引物的5'末端进行荧光标记。PCR体系20μL:10μL 2×Taq plus PCR MasterMix ,1μL 50μg/mL模板DNA、0.3μL 10uM 的正向引物和反向引物、和去离子水补足至20μL。PCR 扩增程序如下:94℃预变性3min;94℃变性30s,53~58℃退火30s,72℃延伸45s,运行32个循环;72℃最终延伸7min,最后4℃保存终止反应。利用毛细管电泳对PCR产物进行分析,使用genemarker 2.0进行结果的确认和读取。
1.4 数据分析处理
使用GenAlex 6.41估算6个种群的遗传参数,包括位点比较等位基因数(Na)、有效等位基因数(Ne)、观测杂合度(Ho)、期望杂合度(He)、shannon多样性指数(I)、群体内固定系数(F)等来衡量6个杜鹃种群内遗传多样性的水平。并计算种群间遗传分化系数(Fst)、基因流(Nm)、Nei’s标准遗传距离、亲缘关系及主成分分析(PCoA)和分子方差分析(AMOVA)等,并根据 Nei’s标准遗传距离,利用 NTSYSpc 2.10e软件非加权配对算术平均法(UPGMA)对各居群进行聚类来分析种群间的亲缘关系。
2 结果与分析
2.1 引物筛选
使用6个不同种的杜鹃花基因组DNA进行了12对EST-SSR引物组合筛选,共筛选出6对电泳条带清楚,多态性高的引物组合用于试验样品的遗传分析。

表1 筛选出的6对EST-SSR引物的序列信息
2.2 6种杜鹃花资源的遗传多样性分析
根据GenAlex分析结果(表2)可知,四明山6种野生杜鹃花的遗传多样性参数差异明显。等位基因数(Na)和有效等位基因数(Ne)为1.75~5.50、1.33~2.87,均表现为马银花最大,华顶杜鹃最小。马银花和华顶杜鹃的观测杂合度(Ho)>期望杂合度(He),其余4种则Ho<He。Shannon’s多样性指数(I)比较得出,马银花和羊踯躅的遗传多样性显著高于华顶杜鹃,排序为马银花>羊踯躅>映山红>云锦杜鹃>满山红>华顶杜鹃。比较F固定指数可知,映山红0.621最高,满山红和云锦杜鹃接近于0,而华顶杜鹃最低为-0.385。
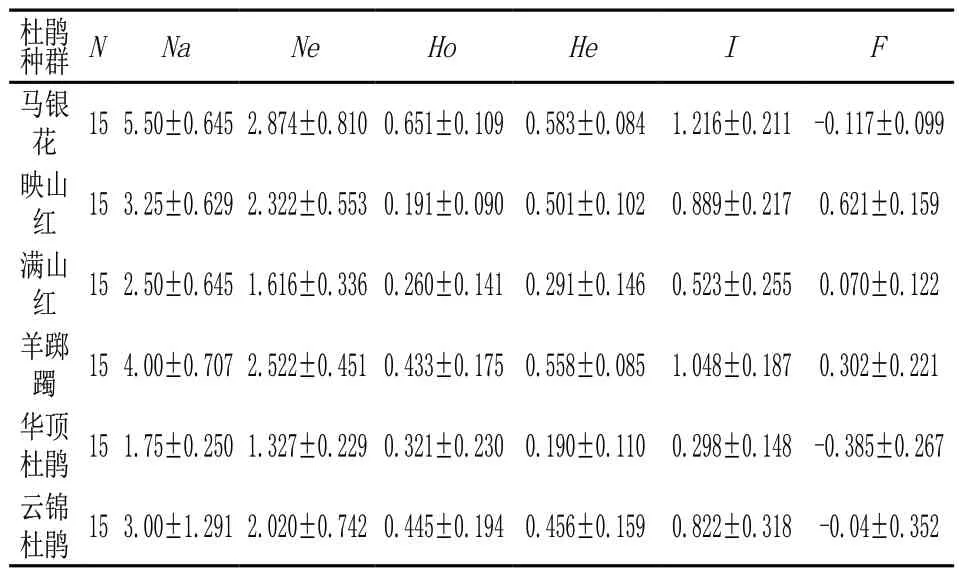
表2 6个杜鹃花属种群遗传多样性指数
由表3可知,四明山野花杜鹃生资源种群间遗传分化系数Fst为0.125~0.420,平均值0.274,马银花与华顶杜鹃种群间的遗传分化最大,与羊踯躅种群间的遗传分化最小;种群间基因流(Nm)为 0.345~1.743,平均值0.786,马银花与羊踯躅种群间的基因流最大,与华顶杜鹃种群间的基因流最小。野生杜鹃种群的分子方差(AMOVA)分析显示(表4),31%的遗传变异存在于种群间,69%的遗传变异存在于种群内。

表3 6个杜鹃花属种群成对分化系数及基因流分析

表4 6个杜鹃花属种群分子方差(AMOVA)分析
2.3 6种杜鹃花资源的亲缘关系分析
由表5可知,6种杜鹃花亲缘关系为0.114~0.816,遗传距离为0.203~2.173。其中马银花和满山红遗传距离最大为2.220,亲缘关系最远为0.109;满山红与华顶杜鹃遗传距离最小为0.252,亲缘关系最近为0.778。利用 Nei's遗传距离,采用平均聚类法对6种杜鹃花自然种群进行聚类分析(图1),6种野生杜鹃花资源明显聚为2组4类,映山红、满山红和华顶杜鹃聚为1类,其他3种单独为1类。利用 GenAlEx 分析种群间主成分分析(图2),第一主成分(PC1)和第二主成分(PC2)的贡献率分别为 66.26%和20.64%,累计贡献率86.90%。图中2个种的距离越近,说明其遗传背景差异越小,亲缘关系越近。结果表明,映山红、满山红和华顶杜鹃明显聚在一起,与其他3种彼此距离较远,此结果和基于Nei's 遗传距离得出的聚类结果一致。
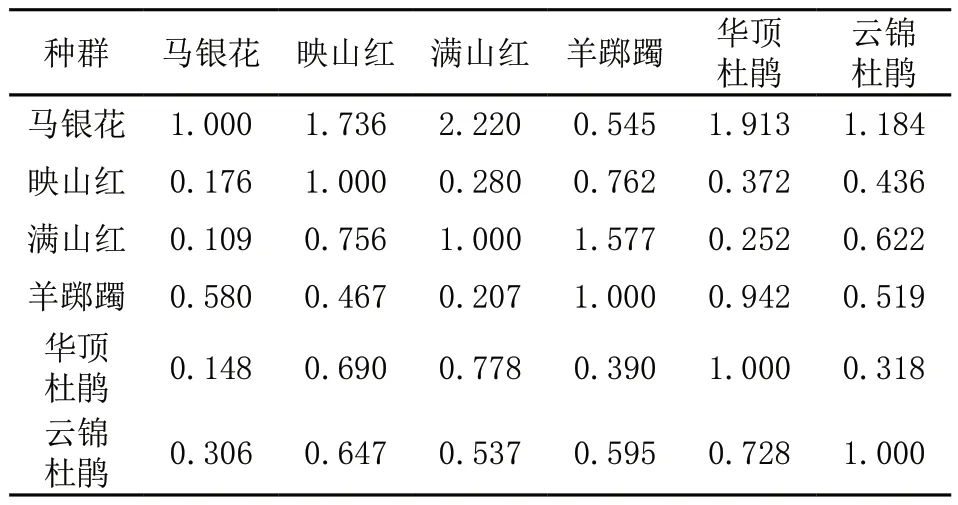
表5 6个杜鹃属种群遗传距离和亲缘关系

图1 6个杜鹃花的聚类分析图
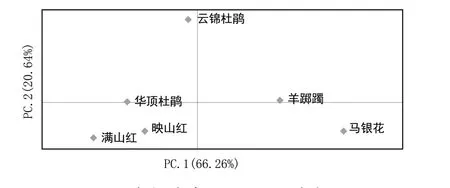
图2 6个杜鹃花种群的PCA分析
3 讨 论
3.1 遗传多样性分析
遗传多样性是衡量一个物种环境适应力和进化潜力的重要指标,是物种生存和进化的内在基础,遗传多样性越高,适应性越强和进化潜力越大[12]。本研究选用了同属植物红马银花的12对EST-SSR引物筛选出了6对扩增效果好、多态性高的引物进行了四明山区域内6种杜鹃花属植物的遗传多样性分析,表明杜鹃花属植物的遗传背景具有很高的相似性,引物的通用性较好,所用的EST-SSR引物可推广应用于更多杜鹃花属植物。结果显示四明山区域内的马银花 (He=0.583,I=1.216)、羊踯躅(He=0.558,I=1.048)、云锦杜鹃(He=0.456,I=0.822)跟同属的其他杜鹃相比拥有很高的遗传多样性,这和相关杜鹃花遗传多样性研究结论基本一致[16,18,19]。调查区域内羊踯躅种群的数量极少,但其遗传多样性却不低,推测造成这种现象的原因是区域内近期花木种植造成的羊踯躅适生环境破坏和因其独特的药用价值带来的大肆采挖导致的居群数量骤减,这和红马银花[18]、白桫椤[20]、栌菊木[21]的致濒因素一致。华顶杜鹃He、I两个遗传多样性参数均为6种杜鹃花资源中最低(He=0.190,I=0.298),表明华顶杜鹃在四明山野生杜鹃花种群中遗传多样性水平最低。再加上其生境岛状狭域分布、野外数量很少、种群结构多为衰退型[22],属于典型的极小种群濒危植物,如果不及时加以保护,其潜在的基因资源就会随植株的消亡而消失。依据Nagylaki[23]的理论,固定指数(F)大于0的映山红和羊踯躅存在较高的近交现象,云锦杜鹃和满山红固定指数接近0以自由交配为主,马银花和华顶杜鹃固定指数小于0,主要以异交为主。
遗传分化系数(Fst)和基因流(Nm)可以用来描述种群间的分化程度,种群间分化系数越大,基因流越小,则种群间差异就越大。四明山野生杜鹃种群间的Fst均值为0.274,大于贵州百里杜鹃(Fst=0.133)[24]和大别山地区杜鹃(Fst=0.175)[25]种群间的遗传分化系数,并且超过0.25,说明四明山区域内杜鹃花资源已存在很大的遗传分化。基因流(Nm)大于1时,种群间的基因交流比较广泛,能够抵制种群间由遗传漂变引起的遗传分化,而基因流小于1,则种群间基因交流受阻,遗传分化加重[26]。四明山野生杜鹃花种群间基因流均值0.786,表明种群间基因交流受阻。本研究中华顶杜鹃与马银花的遗传分化系数最大、基因流最小,表明该区域内两种杜鹃花的差异最大。6种杜鹃自然种群的分子方差(AMOVA)分析显示,31%的遗传变异存在于种群间,69%的遗传变异存在于种群内。表明种群间已出现明显分化,该结果与种群间遗传分化系数得出的结论相一致。
3.2 亲缘关系分析
四明山6种野生杜鹃花资源隶属于4种杜鹃花亚属,其中云锦杜鹃属于常绿杜鹃亚属(Subgen.Hymenanthes(Blume)K.Koch),马银花属于马银花亚属(Subgen.AzaleastrumPlanch.ex K.Koch),羊踯躅属于羊踯躅亚属(Subgen.Pentanthera(G.Don) Pojarkova),映山红和满山红属于映山红亚属(Subgen.Tsutsusi(G.Don)Pojarkova)。本研究聚类结果和PCA分析结果一致,均表现为映山红和满山红具有很近的亲缘关系,符合经典系统分类;结果还显示马银花和羊踯躅也有较近的亲缘关系,这和李美芹[16]、张哲玮[27]、黄承玲[28]的聚类结果基本一致;以上都表明了本研究得出的亲缘关系具有较好的准确性。华顶杜鹃为丁炳扬1990年才发现的新种[29],根据其落叶习性、小枝具柔毛和叶3~5枚假轮生状密集枝顶,将其归入映山红亚属轮叶杜鹃组,但其种子形态具有许多独特之处,与其余种类均明显不同[30]。后来章月皎以花芽、叶芽、花粉以及种子形态特征为依据认为华顶杜鹃应该属于羊踯躅亚属的成员[14]。颜士辉等做了华顶杜鹃ISSR反应体系的优化及亲缘关系的初步分析,结果对华顶杜鹃的分类地位没有得到明确结论[15]。李美芹等以宁波地区的杜鹃花种质资源为研究材料开发EST-SSR引物并探究遗传多样性及遗传结构,聚类结果显示华顶杜鹃和云锦杜鹃聚为一类[16],但华顶杜鹃和云锦杜鹃植株形态和特性均差异很大,很难划归于同一类群。本研究通过筛选的ESTSSR引物对华顶杜鹃的遗传多样性和亲缘关系做了分析,结果显示华顶杜鹃与满山红亲缘关系最近,与羊踯躅和马银花亲缘关系较远,本结论与丁炳扬最初根据其形态特征作出的分类归属一致,佐证了华顶杜鹃隶属于映山红亚属轮生叶杜鹃组的结论。但结论仍然受到样本量的代表性和ESTSSR分子标记的引物以及方法局限性的限制,后续可利用其他分子标记或者基因测序等方法进一步验证。
优先保护濒危植物有助于延缓物种多样性的流失、维护生态平衡以及促进生态可持续发展,对我国生物多样性保护具有重要意义[31]。羊踯躅具有很高的药用价值,花、果实和根等均可供药用,可祛风除湿、舒筋活血、镇痛止痛。虽然试验结果显示其仍具有较高的遗传多样性水平,但因人为掠夺式挖掘,导致区域内植株急剧减少,建议采取抢救式种质资源异地保存,辅以种苗扩繁的资源保护措施。华顶杜鹃具有众多特异的植物学性状,在杜鹃花属植物的进化和系统发育中具有重要的研究价值,其种群数量极少、遗传多样性水平低,属于典型的极小种群濒危植物。建议在就地保护和迁地保育的基础上,开展人工多种源种苗繁育并尝试野外回归工作,增加野外种群的植株数量、优化种群的年龄结构、丰富种群的遗传多样性,缓解其濒危程度。
