云南松苗木生物量分配与个体大小关系研究
2023-01-18蔡年辉唐军荣许玉兰
王 瑜,蔡年辉,陈 林,唐军荣,许玉兰,陈 诗
(1.西南林业大学 西南地区生物多样性保育国家林业局重点实验室,昆明 650224;2.西南林业大学 西南山地森林资源保育与利用教育部重点实验室,昆明 650224)
植物的生物量分配能体现其对环境的响应与适应性特征,研究生物量的分配是植物净碳获取的综合结果与重要驱动因素,对植物的生长发育有重要影响,同时可反映植物生长资源利用策略[1-3],因此探讨其分配特征受到广泛关注[2-6]。根、茎、叶是植物体基本的形态学单位,根主要是吸收生长环境的养分、水分[1,7],茎起到输导水分、养分功能及机械支撑与固定作用[1,8-9],而叶片是重要的光合器官,其碳同化产物是植株生物量积累和产出的生物学基础[1],各器官分配格局的可塑性可表示生物量分配格局对环境的响应模式[10],生物量分配模式影响着植物个体生长和繁殖甚至到整个群落的质量和能量流动等功能[8],不仅有助于从易测量的地上生物量估算地下生物量,还体现着植物不同的生态策略[11],反映出光合产物对植物不同器官(地上不同器官之间,地上器官和地下器官之间)的投资以适应环境条件[12-14]。植株生物量分配受多因素的影响,包括材料起源[15-17]、年龄[1,17]、个体大小、生境条件[2-3,20]等,目前主要有生物量异速生长理论和生物量最优分配理论解释,生物量异速生长理论认为,生物量的分配受个体大小的影响[21-22],其分配比例会随植株的大小变化而变化[3,9,23]。生物量最优分配理论认为,植株会将更多的生物量分配到能获得限制性资源的器官中,以维持最大的生长速率[24-26]。
云南松是我国西南地区的主要用材树种和生态树种,目前,针对云南松苗木生物量在种源[15]、家系[16,27]、林龄[17]、干型[28]及其苗木分化[19]等方面开展研究,同时也探讨了苗木各器官生物量积累的异速生长关系[15-16,29]。但是,这些研究针对各器官之间的生物量分配方面较少,对于生物量分配格局随苗木个体大小变化规律方面尚未见报道。研究表明,植株生长过程中,可能随个体大小或发育阶段的不同而生物量分配特征有所差异,这种差异可反映苗木的生长对策[3]。本文以云南松苗木为材料,通过整株收获法测定云南松苗木根、茎、叶各器官的生物量,分析云南松苗木生物量分配特征及其随苗木个体大小变化规律,探究其资源分配策略的变异性和保守性,以期为综合揭示云南松苗木生物量特征,提高云南松苗木生物量及其碳储量的估算精确度提供参考,这对开展云南松定向培育也具有重要的现实意义。
1 材料与方法
1.1 研究材料
在云南省昆明市宜良县进行球果采集,将采摘的球果带回实验室晾晒风干,待球果开裂取出球果中的种子,以5cm×10cm的株行距播种于西南林业大学格林温室内,定期对云南松进行浇水、喷药等苗期养护管理。
1.2 指标测定
对2a生苗木,采用整株收获法将云南松个体整株挖出[18],共615株,编号后带回实验室,用直尺和游标卡尺分别测量各株的苗高(精确至0.01cm)、地径(精确至0.01mm),然后用剪刀将根系从根茎处剪下[30],将地上部分茎、叶以及地下部分即根分开,其中地下部分用清水洗净并沥干[31],称量其鲜质量,分别装入标记好的纸袋中,在105℃的烘箱中杀青30min后,调至80℃进行烘干处理至质量恒定,测量根、茎、叶各构件的干质量,即为生物量,精确至0.001g。并计算相关指标:地上部分生物量=茎生物量+叶生物量,非光合器官生物量=茎生物量+根生物量,单株生物量=根生物量+茎生物量+叶生物量,生物量分配比=各构件生物量÷单株生物量×100%,叶茎比=叶生物量÷茎生物量,叶根比=叶生物量÷根生物量,茎根比=茎生物量÷根生物量,地上部分与根比=地上部分生物量÷根生物量,叶与非光合器官比=叶生物量÷非光合器官生物量,高径比=苗高÷地径,苗木质量指数=单株生物量÷(高径比+地上部分生物量÷根生物量)[32-33]。
1.3 数据分析
采用SPSS 17.0进行描述统计分析,其中变异系数=标准差/平均值×100%。同时对苗木个体大小(苗高、地径、苗木质量指数)与各构件生物量分配间的相关关系进行分析(Pearson,p=0.05),利用幂函数构建生物量分配与个体大小间的曲线方程[3,5-6],采用Excel 2010作图。
2 结果与分析
2.1 苗木生物量及其分配格局
对云南松苗木实测的生物量进行分析,比较根、茎、叶、地上部分和非光合器官各构件在单株水平上的分配比,以及各构件之间的分配比。由表1可知,云南松苗木根、茎、叶生物量分别占单株生物量的15.57%,26.30%和58.13%,即叶生物量分配比>茎生物量分配比>根生物量分配比,组合后的构件地上部分生物量和非光合器官生物量分别占单株生物量的84.43%,41.87%,光合器官(叶)生物量>非光合器官(茎+根)生物量,地上部分生物量分配比远远大于地下部分生物量分配比。但生物量分配比的变异幅度较大,根、茎、叶、地上部分、非光合器官生物量分配比分别波动于 6.08%~48.77%,2.81%~71.46%,13.51%~79.92%,51.23%~93.92%,20.08%~86.49%。从生物量分配比的变异系数来看,单株水平上以根分配比的变异数最大(32.91%),以地上部分分配比的变异系数最小(6.07%)。各构件生物量分配格局的变异系数波动于37.34%~60.47%,以地上部分与根比的变异系数低,叶茎比的变异系数高。综合来看,云南松苗木生物量在不同构件间的分配存在差异性。
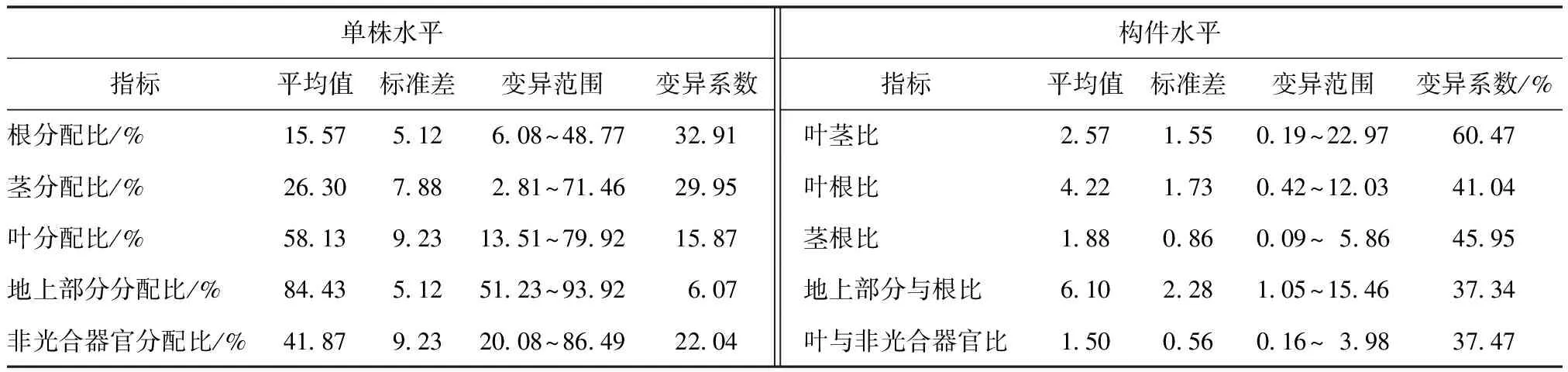
表1 云南松苗木生物量分配Tab.1 Biomass allocation of Pinus yunnanensis seedling
2.2 苗木各构件之间生物量分配关系
将上述各构件两两组合进行线性关系拟合,分析云南松苗木生物量各构件之间生物量分配关系,结果表明,各构件间均具有很好的线性关系(p<0.01)。从图1中可以看出,各方程的斜率差异较大,波动于0.871 9~4.475 6,以地上部分与根之间拟合方程的斜率最大,而叶与非光合器官之间拟合方程的斜率最小。
2.3 苗木生物量分配随个体大小变化规律
利用相关性分析各构件生物量分配与苗木大小,即苗高、地径、苗木质量指数间的相关性,结果如表2所示。云南松苗木根、茎、叶、地上部分、非光合器官各构件生物量分配比与苗木大小,即苗高、地径、苗木质量指数间的相关性有所不同,其中苗高、地径、苗木质量指数与茎分配比、非光合器官分配比间均呈极显著正相关关系(p<0.01),与叶分配比均呈极显著负相关关系(p<0.01)。苗高与根分配比为极显著负相关关系(p<0.01),与地上部分分配比为极显著正相关关系(p<0.01);地径与根分配比、地上部分分配比间的相关性均不显著(p>0.05);苗木质量指数与根分配比为显著正相关关系(p<0.05)、与地上部分分配比为显著负相关关系(p<0.05)。与生物量分配格局包括叶茎比、叶根比、茎根比、地上部分与根比、叶与非光合器官比之间也存在不一样的相关性,其中叶茎比、叶与非光合器官比和苗高、地径、苗木质量指数间均为极显著负相关关系(p<0.01)。茎根比与苗高、地径均为极显著正相关(p<0.01),与苗木质量指数间为显著正相关(p<0.05)。叶根比与苗高、地径和苗木质量指数间均为负相关关系,其中与地径和苗木质量指数的相关性达极显著水平(p<0.01)。地上部分与根比与苗高间呈极显著正相关关系(p<0.01),与地径呈显著负相关关系(p<0.05),与苗木质量指数间为极显著负相关关系(p<0.01)。综合来看,苗木生物量分配与苗木个体大小间存在较为密切的关系。
图2所示,云南松苗木生物量分配(包括叶茎比、茎根比、叶根比、地上部分与根比、叶与非光合器官比)与苗高、地径间均有较好的幂函数关系。从拟合曲线可知,苗木高生长或粗生长较小时,将生物量更多的分配给叶,表现为叶茎比、叶根比、叶与非光合器官比较高,即呈现负相关关系。非光合器官即茎、根的分配来看,苗木高生长或粗生长较小时,生物量更多的分配给地下部分的根,表现为茎根比小。从地上部分器官(茎、叶)与地下部分器官(根)来看,苗高、地径与其相关性不一致,其中与苗高间为正相关,与地径间为负相关,即苗木较高或较细时,生物量向地上部分分配较多。综合来看,苗木个体小时,表现为生物量倾向于地上部分的分配,其中又以向叶的分配占优势,茎的分配相对减少,苗木粗生长受影响,即与地径间呈负相关关系。
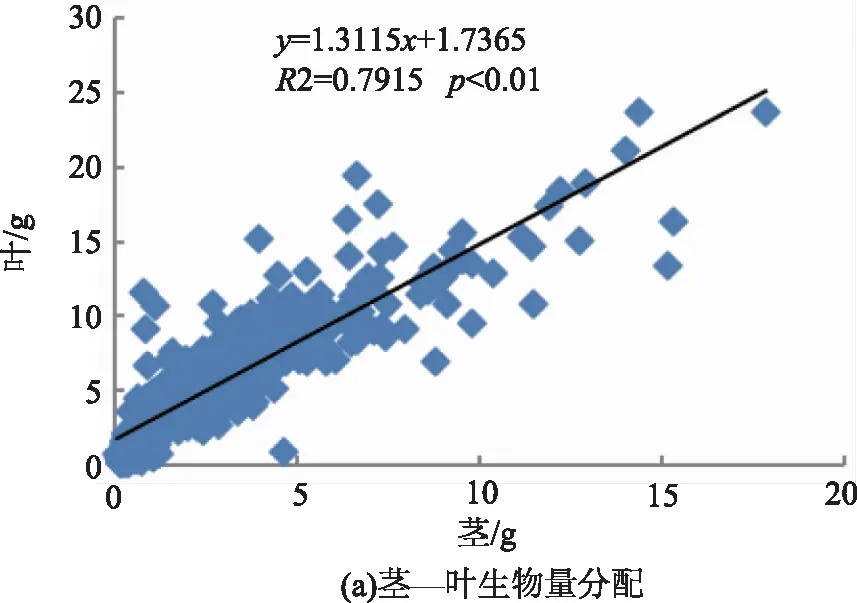

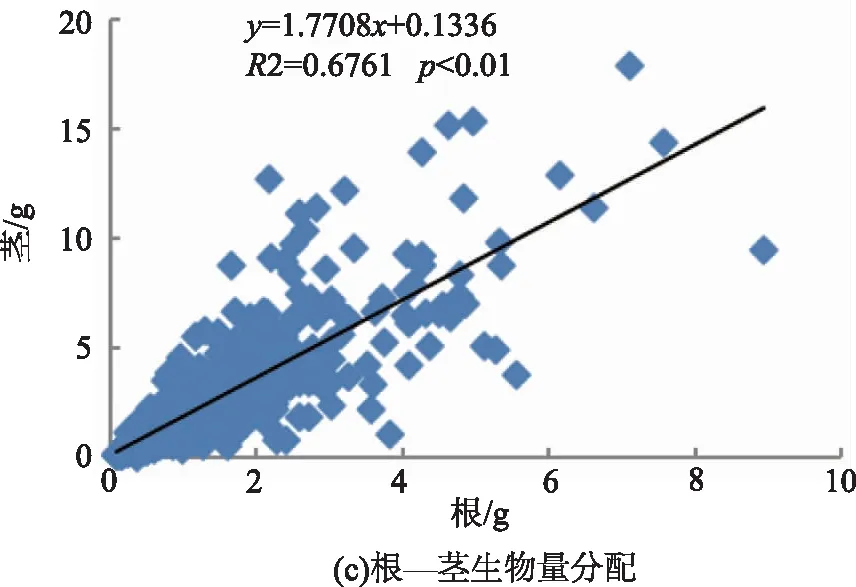


图1 云南松苗木各构件间生物量分配关系Fig.1 Biomass allocation among components of P. yunnanensis seedlings

表2 云南松苗木生物量分配与个体大小的相关性分析Tab.2 Correlation analysis of biomass allocation and individual size in P.yunnanensis seedlings

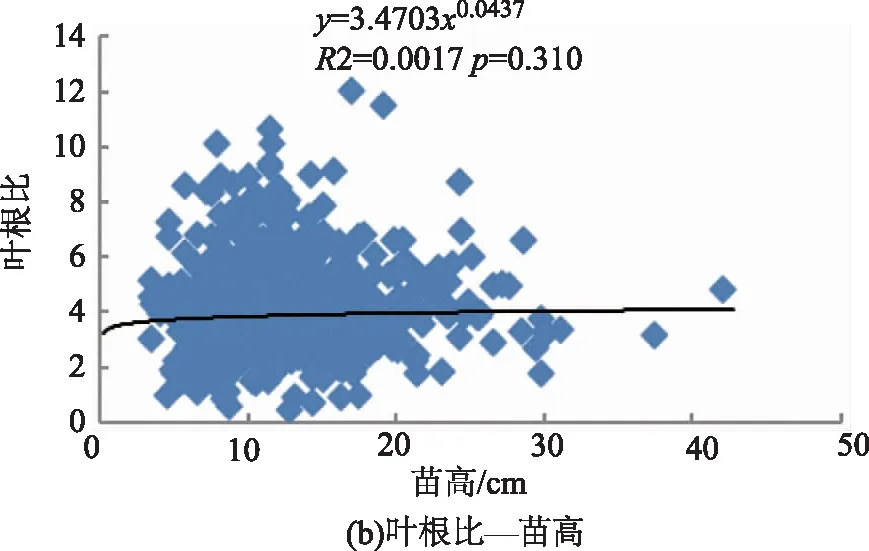



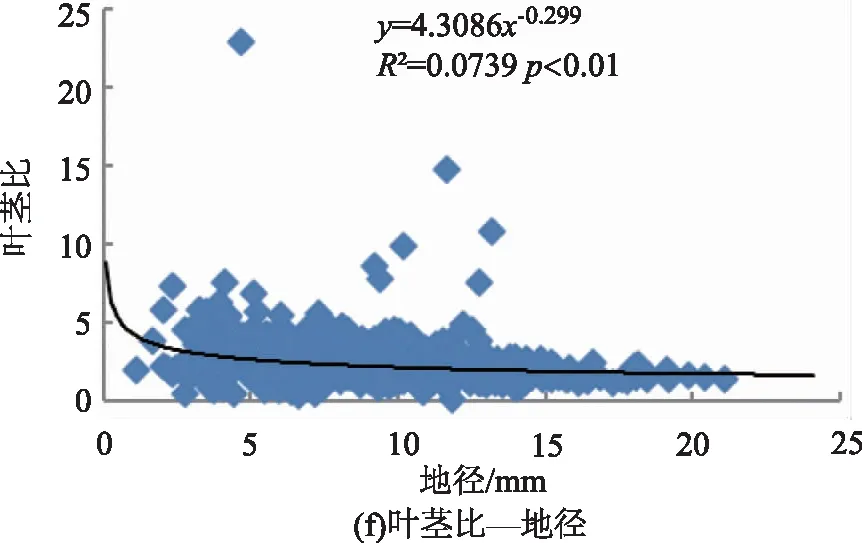
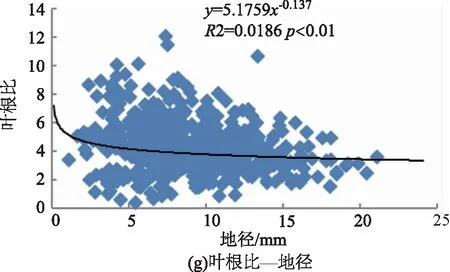



图2 云南松苗木生物量分配与个体大小间的关系Fig.2 Relationship between biomass allocation and individual size in P.yunnanensis seedlings
3 结论
1) 生物量分配可反映植株生长的生态对策[2-3,11],以根、茎、叶3个器官来看,云南松苗木生物量分配表现为叶>茎>根,地上部分生物量分配远远大于地下部分即根生物量分配,光合器官即叶生物量分配也大于非光合器官。由此表明,云南松苗期阶段以叶生物量分配为主,这样的分配可增强光合作用,积累有机物质,利于苗木生长,即植物个体较小时,更注重叶资源的分配,以最大化光获取能力[24],维持最大的生长速率[18],体现生物量的最优分配理论。但同时也看到,云南松苗木生物量分变异系数较大,即苗木生物量分配具有较大的变异性,这在其它树种中也有研究报道[34],这种变异性一方面可增加苗木的适应能力,同时也为苗木生物量的评价与精确估算带来挑战。
2) 前期的研究表明,苗木生长量与生物量之间存在一定的相关性,可利用苗木的生长量如苗高、地径等去估测苗木生物量[35-36]。本研究分析可知,不同构件间生物量也存在较为密切的关系,拟合的线性方程效果较好。因此,可利用易测构件估算不易测定构件的生物量[34],从而为生物量的快速估算提供途径。生物量分配随着个体大小变化将资源同步按一定的比例分配到各构件,实现不同构件在不同个体大小下的比例均衡性,这种分配的稳定性也反映了云南松苗木生物量分配存在权衡[25]。
3) 本研究以苗高、地径及其综合指标苗木质量指数表示苗木个体大小,分析了生物量分配与个体大小间的关系,结果表明,生物量分配与个体大小间关系显著(p<0.05),表明生物量分配随个体大小发生变化,即生物量分配受个体大小的影响,在一定程度上体现了生物量分配的异速生长理论,类似研究也有报道,即苗木生物量分配存在大小依赖性[6,34]。其中地上部分与根比和苗木质量指数、地径间呈极显著负相关关系,表明苗木较小时地上部分的分配相对增加。进一步分析揭示叶茎比、叶根比、叶与非光合器官和苗木个体大小间均呈负相关关系,且基本达到极显著水平(p<0.01),拟合的幂函数曲线方程也表明苗木较小时,生物量倾向于地上部分尤其向叶的分配,即个体较少时,倾向于叶的分配,体现生物量的最优分配理论[4]。
4) 在苗木相同个体大小的情况下,生物量分配也具有较大的变异性。因此,很难用统一模式分析生物量分配特征,这可能与生物量分配受多种因素的影响有关。如,孟欢欢等[6]研究表明,草本植物层片根冠比与栽植密度间呈线性正相关关系,而去除层片内的类短命植物囊果苔草后,根冠比与栽植密度间呈显著负相关关系;孙迎涛等[2]研究表明,不同沙漠化程度(潜在、轻度、中度、重度)下油蒿个体的生长及生物量分配格局不同,会随着沙漠化程度的加剧发生调整与权衡,由以营养生长为主向生殖生长为主发生转变。这些研究也进一步证实苗木生物量分配会受到外界环境及个体发育的影响,并呈现明显的可塑性,在一定程度上反映资源分配的保守性与可塑性的权衡[3],植物通过改变个体的形态特征以及调整构件间生物量分配适应不同环境[5]。生物量分配的2种理论即最优分配理论和异速生长理论可以相互补充[34],以更加全面地解释植物生物量分配特征。当然,这种关系是会随苗木生长发育阶段或环境条件改变而发生变化,值得进一步研究。
