Rac1突变重组体的构建及对JAK/STAT信号通路的影响
2023-01-16尚娇娇叶守东
尚娇娇,胡 玲,方 健,叶守东,孙 敏
(安徽大学 生命科学学院,安徽 合肥 230601)
糖尿病肾病(diabetic nephropathy,简称DN)是糖尿病患者的常见微血管并发症之一,其主要病理变化为细胞外基质增加、肾小球肥大,肾小球和肾小管基底膜增厚等,最终导致肾小球硬化伴有肾间质纤维化[1].Ras-related C3肉毒毒素底物1(Ras-related C3 botulinum toxin substrate 1,简称Rac1)是Rho家族小GTPases的一员,被证明参与DN肾小球系膜细胞外基质(extracelluar matrix,简称ECM)聚集[2],高血糖、糖基化终末产物(advanced glycated end products,简称AGEs)、血管紧张素II(Angiotensin II,简称ANG II)、生长因子等水平升高都能引起Rac1过度激活[3-5].Janus激酶/信号转导子和转录激活子(Janus kinase/Signal transducer and activator of transcription,简称JAK/STAT)通路是一条由细胞因子激活的信号通路,在DN中被激活,从而产生转化生长因子[6].转化生长因子β(transforming growth factor-β,简称TGF-β)是一种多功能细胞因子,与增殖、凋亡、分化和纤维化有关,TGF-β作为肾小球硬化进展的中心介质,导致肾脏中异常ECM蛋白的积累和平滑肌α-肌动蛋白的不恰当表达[7-9].由于Rac1和JAK/STAT信号均涉及ECM的形成,因此,Rac1与JAK/STAT信号通路的关系,及在系膜细胞损伤中的作用有待于深入研究,笔者通过构建持续激活型Rac1(G12V)肾小球系膜细胞,研究Rac1激活对JAK/STAT信号通路的影响,及在系膜细胞损伤中的作用.
1 材 料
1.1 试 剂
PiggyBac(PB-FLAG)转座子质粒载体由安徽大学生命科学学院叶守东老师实验室提供;Trizol,Transgene公司;逆转录试剂盒,白鲨生物公司;PCR反应体系,南京诺维赞公司;限制性内切酶BgI II,Xho I,NEB公司;T4 DNA连接酶、DH5α大肠杆菌,TaKaRa公司;DNA凝胶回收试剂盒,杭州倍沃医学科技公司;DNA纯化回收试剂盒,北京天根公司;琼脂糖、DCFH-DA,美国Sigma公司;LB培养基,上海生工公司;引物设计,NCBI数据库;引物合成和测序,安徽通用公司;DNA-Hieff TransTM脂质体核酸转染试和嘌呤霉素,上海翌圣公司;小鼠肾小球系膜细胞株(mouse mesangial cell, 简称MES),中国科学院典型培养物集存库;DMEM,北京索莱宝公司;FBS,杭州四季青公司;NADPH氧化酶活性试剂盒,北京百奥莱博公司;小鼠TGF-β Elisa试剂盒,武汉亚科因公司;FLAG抗体、JAK2抗体、FN抗体、GAPDH抗体,武汉Proteintech公司;p-JAK2抗体,武汉ABclonal公司;STAT3抗体、p-STAT3抗体,美国Cell Signaling Technology公司;HRP二抗,上海萨博公司;羊抗兔FITC荧光二抗,武汉博士德生物公司;硝酸纤维素膜(nitrocellulose filter membrane,简称NC膜),美国GE公司;ECL显影液,上海天能科技有限公司.
1.2 仪 器
PCR仪、NanoDrop超微量分光光度计,美国赛默飞公司;琼脂糖水平电泳仪,北京六一公司;超净工作台,新加坡ESCO;二氧化碳培养箱,日本松下公司;倒置显微镜,日本奥林巴斯公司;多功能酶标仪,美国BioTek公司;JS-1070P型化学发光成像系统,上海培清科技有限公司;DYY-4C型电转膜仪及电泳仪装置,上海天能科技有限公司;徕卡TCS SP8 MP多光子共聚焦显微镜,德国徕卡公司.
2 方 法
2.1 Rac1过表达突变体转移载体质粒的构建
Trizol法抽提系膜细胞全基因组,逆转录为cDNA,由Rac1序列合成引物
GAAGATCTATGCAGGCCATCAAGTGTGTGGTG(上游引物),
CCGCTCGAGTTACAACAGCAGGCATTTTCTCTTC(下游引物)
扩增Rac1目的基因,再以Rac1目的基因为模板,用上游引物
GAAGATCTATGCAGGCCATCAAGTGTGTGGTG,
下游引物
GAAGATCTATGCAGGCCATCAAGTGTGTGGTGGTGGGAGACGTAGCTGTTGGT
扩增出含有上游酶切位点Bgl II及下游酶切位点Xho I的突变体Rac1(G12V)目的基因.将突变体Rac1(G12V)目的基因及带有FLAG标记的PiggyBac(PB-FLAG)转座子质粒载体用限制性内切酶Bgl II和Xho I双酶切,再经胶回收试剂盒回收大小片段,连接酶试剂盒连接大小片段,构建突变重组载体PB-Rac1(G12V).重组质粒转化入DH5α感受态细胞,筛选重组质粒的阳性克隆,扩增后抽提质粒,双酶切PCR鉴定及质粒测序鉴定.
2.2 转染MES细胞
将含10% FBS的DMEM培养基培养的MES细胞,以3×105mL-1密度接种于6孔板中,放置于37 ℃,5% CO2细胞培养箱中培养,待细胞达到60%汇合时开始转染.设置空白对照组、PB空载组、PB-Rac1(G12V)组.DNA-Hieff TransTM脂质体核酸转染试剂复合物的配置:先取250 μL DMEM稀释质粒DNA并混匀;再取250 μL DMEM稀释脂质体核酸转染试剂,室温孵育3 min;再将稀释好的DNA和稀释的脂质体核酸转染试剂轻轻混匀成共500 μL的混合液,并在室温下避光孵育20 min;最后,再加入700 μL的无血清DMEM,使其成为总体积1 200 μL的转染试剂复合物.弃掉MES细胞的培养液,加入转染试剂复合物进行转染,6 h后弃去培养基每孔加入2 mL新鲜的培养基.转染48 h后更换为含终浓度2 μg·mL-1嘌呤霉素的DMEM完全培养基进行筛选.筛选完成后收集细胞样品,Western blot法对细胞进行FLAG标记验证.
2.3 活性氧(reactive oxygen species,简称ROS)及NADPH氧化酶活性测定
正常MES细胞、Rac1(G12V)组MES细胞用DMEM完全培养基混悬后,以3×104mL-1密度分别接种于96孔板和24孔板,每组设置3个复孔,分别用于检测ROS、NADPH氧化酶.将细胞培养于37 ℃,5% CO2培养箱中,48 h后弃去96孔板培养基,预冷PBS清洗2遍,每孔加10 μmol·L-1DCFH-DA 100 μL,37 ℃避光孵育30 min,PBS洗3遍,酶标仪激发光485 nm,发射光530 nm检测荧光值.
48 h后弃去24孔板培养基,按照NADPH氧化酶试剂盒说明书检测NADPH氧化酶活性.
2.4 Elisa检测TGF-β水平
正常MES细胞、Rac1(G12V)组MES细胞以3×104mL-1密度接种于24孔板,每孔1 mL,每组设置3个复孔.48 h后收集正常组、Rac1过表达突变组的MES细胞上清于4 ℃,12 000 r·min-1离心5 min,取上清,按照试剂盒说明书检测TGF-β的表达含量.
2.5 Western blot检测
正常组、Rac1(G12V)组MES细胞以3×105mL-1密度接种于T25细胞瓶中,48 h后收集细胞于离心管中,离心弃上清,每管加入适量预冷的RIPA裂解液,超声裂解2次,冰上静置1 h后,于4 ℃,12 000 r·min-1离心20 min.离心结束后取上清进行BCA蛋白定量测定,调蛋白浓度一致后加入上样缓冲液混匀并在沸水浴中煮5 min,制成的样品于-20 ℃冰箱保存.样品进行SDS-PAGE电泳、转膜、封闭,一抗4 ℃孵育过夜,二抗室温孵育2 h,TBST清洗后ECL曝光,并使用ImageJ进行灰度值分析.
2.6 免疫荧光
正常MES细胞、Rac1(G12V)组MES细胞以3×104mL-1密度接种于激光共聚焦专用细胞培养皿中,每孔2 mL,48 h后取出培养皿,弃上清,预冷PBS 清洗2遍,冰乙醇室温固定20 min,PBS洗3遍,加入5% BSA 37 ℃封闭1 h,弃BSA,一抗4 ℃孵育过夜,PBS洗3遍,加入FITC标记二抗,37 ℃避光孵育1 h,DAPI孵育15 min,PBS洗3遍.每个培养皿中加入1 mL PBS覆盖细胞,置于徕卡TCS SP8 MP多光子共聚焦显微镜下观察并拍照.
2.7 统计学分析

3 结 果
3.1 成功构建Rac1(G12V)持续磷酸化突变体并在MES细胞中表达
该实验构建了Rac1点突变体(G12V),图1(a)表示PB-Rac1(G12V)质粒中Rac1(G12V)的琼脂糖电泳图,图中Rac1(G12V)的条带在正确位置,重组构建成功.序列比对结果显示,全自动测序的结果中,Rac1(G12V)与GeneBank中Rac1序列仅在CDS区第35位碱基处存在差异(A-T),其余序列完全一致.证明所构建的Rac1持续磷酸化突变体G12V序列均完全正确.转染成功后收集细胞,制备样品以抗FLAG的抗体进行免疫印迹检测Rac1突变体的蛋白表达,结果显示(图1(b)~(c)),Rac1突变体在转染的MES细胞中得到很好的表达,细胞形态正常.因此,笔者成功构建的Rac1持续磷酸化突变体可用于Rac1功能调节的进一步研究.
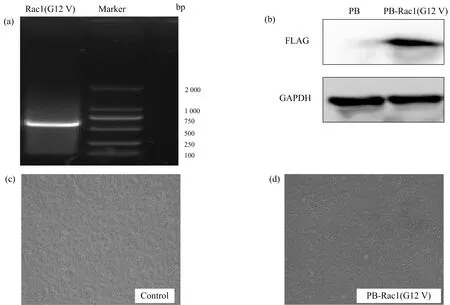
(a)Rac1(G12V)的琼脂糖电泳图;(b)转染的MES细胞中Rac1突变体蛋白表达的免疫印迹检测;(c)~(d)转染的MES细胞.
3.2 Rac1(G12V)对MES中NADPH氧化酶和活性氧ROS的影响
NADPH氧化酶是一种多亚基复合体,是细胞中产生ROS的主要来源,Rac1是其组成亚基之一.Rac1活化可导致NADPH氧化酶活性增加,因此观察Rac1持续激活对NADPH氧化酶活性和产生ROS的影响,结果如图2所示.

与对照组相比,
图2显示,Rac1(G12V)组的NADPH氧化酶活性明显高于对照组,其ROS的表达水平也明显高于对照组.提示Rac1活化导致NADPH氧化酶活性增加并产生大量ROS.
3.3 Rac1(G12V)对MES中JAK2/STAT3信号通路的作用
3.3.1 Rac1(G12V)激活JAK2/STAT3信号通路
JAK2/STAT3信号通路在DN状态下被各种因素,如高糖、AGE、氧化应激等激活,进一步探究Rac1激活对JA2/STAT3信号通路的影响,结果如图3所示.

(a)p-JAK2/JAK2的表达,(b)p-STAT3/STAT3的表达; 与对照组相比,
由图 3可知,Rac1(G12V)组p-JAK2/JAK2以及p-STAT3/STAT3的表达比对照组显著增加,提示Rac1能激活JAK2/STAT3信号通路.
3.3.2 Rac1(G12V)对MES产生TGF-β的作用
为进一步验证Rac1对JAK2/STAT3信号通路的激活作用,检测其下游信号分子TGF-β的表达,结果如图4所示.

与对照组相比,
图4显示,Rac1(G12V)组的TGF-β表达水平明显高于对照组,提示Rac1活化可以激活JAK2/STAT3信号通路.
3.3.3 Rac1(G12V)对MES纤连蛋白(FN)的影响
TGF-β被认为可以促进系膜细胞肥大,促使ECM 积聚,从而在DN肾小球硬化和肾间质纤维化中起着重要作用,而纤连蛋白(fibronectin,简称FN)是ECM的主要成分,因此通过检测FN水平来反应ECM积聚情况,结果如图5所示.
由图5可知,Rac1(G12V)组的FN表达水平明显高于对照组,免疫荧光实验显示Rac1(G12V)的FN表达增强,提示突变过表达的Rac1能增加系膜细胞ECM的积聚.
4 讨 论
Rho(小G蛋白)是Ras蛋白超家族的一个亚群,具有GTP酶活性,又称为Rho GTP酶(GTPase),在该家族中,Rac1是研究最广泛的成员之一,于1989年被发现为Ras相关的C3肉毒毒素底物1,分子量为21 kDa,由192个氨基酸编码组成[10].随后,它被证明在多种细胞中发挥基本作用,包括肌动蛋白细胞骨架重组、细胞转化、诱导DNA合成、超氧化物产生和细胞迁移[11].Rac和Rho GTPase是GTP结合蛋白,起分子开关的作用.当与GTP结合时,GTPase可以与效应分子或靶标分子相互作用并启动信号转导,当它们与GDP结合在一起时,这种信号传导就被抑制[12].Rac1的异常表达或激活可引起异常的细胞信号传导,从而导致病理损伤,Rac1蛋白作为NADPH氧化酶的一个活性亚基,其活性变化将会直接影响其所依赖的Rac1/NADPH 信号通路的激活和抑制,而来源于NADPH氧化酶的ROS被认为是糖尿病及其并发症发病的始作俑者[13].此外,GTPases的Rac/Rho家族是肌动蛋白细胞骨架的关键调控因子[14],在DN肾纤维化中形成的作用已被证实,Hubchak等[15]认为Rac1通过一种不直接改变经典Smad3信号事件的机制来增强TGF-β1诱导的I型胶原表达.
JAK/STAT信号通路是一条具有信号转导和基因转录双重功能的信号通路,可介导炎症、细胞增殖和纤维化信号的传递[16].在DN中,JAK2/STAT3信号通路被高血糖、氧化应激、AGE等激活,引起TGF-β过表达,导致ECM扩张、肾小球基底膜以及纤连蛋白FN、层粘连蛋白和I胶原蛋白的积累,最终加速DN的发生和发展[17].此外,研究表明Rac1突变体显著提高了JAK2的活性,突变激活的Rac1增加JAKs的磷酸化和STATs的转录活性[18].Rac1可以通过激活 IL-6/JAK2通路或直接与STAT3结合来激活STAT3和信号转导[19].
为了研究Rac1激活在系膜细胞损伤中的作用,笔者构建了持续激活的Rac1(G12V)重组载体,导入MES细胞后稳定表达.G蛋白偶联受体不仅能在激动剂刺激后传递信号,还能自发偶联到信号转导通路,即在没有激动剂存在的情况下本身可以转变成活性构象,传递信号[20].笔者所设计的 Rac1 组成型激活突变体 Rac1(G12V)是对照 Rac1的组成型突变体将P-loop上12位的G/甘氨酸突变为V/缬氨酸,能够模拟野生型G蛋白的激活状态,人为构建组成型激活突变体[21].研究结果表明,由持续性Rac1激活可激活JAK2/STAT3信号通路,使TGF-β过度表达及细胞外基质积累.但是Rac1激活是通过ROS还是直接激活STAT3有待于进一步研究证实.
