物种和功能维度上贵州茂兰藓类植物多样性格局及其环境因素
2023-01-14黄若玲李丹丹吕金桥张永昌郭水良
黄若玲,李丹丹,吕金桥,张永昌,杨 军,邓 坦,何 林*,郭水良*
(1.上海师范大学生命科学学院,上海 200234;2.遵义师范学院生物与农业科技学院,贵州遵义 563006)
0 引言
喀斯特地貌在我国西南地区分布广泛,在其上发育的喀斯特森林生境复杂,具有丰富的生物多样性,对于稳定其地区生态环境有积极作用.茂兰国家级自然保护区具有典型的喀斯特地貌,有漏斗、沟谷、盆地等景观.该保护区拥有同一纬度上保存下来的面积最大、比较集中、原生性强、较为稳定的亚热带喀斯特森林生态系统.由于该地区特有的石灰岩地质环境,及复杂的生境类型,使其植物种类组成、植物区系成分及群落结构十分复杂[1-2].
苔藓植物是种数上仅次于被子植物的特殊植物类群,形体小、枝叶交错卷曲,多数呈丛生状、匍匐状或垫状,有吸水快、蓄水量大的特点[3].喀斯特地区的苔藓植物中存在相当多附着于岩石表面生长的种类,能够改善其下垫面的水热条件,为维管植物提供生境基础.石生苔藓分泌的酸性物质能够溶蚀岩石表面,促进母岩的风化及成土过程[4].与此同时,苔藓植物特殊的生长形式能够将径流和空气中细小的土壤颗粒拦截并沉积于假根下,起到良好的固土作用,为其他植物的生长提供了一定的基质[5].
喀斯特石漠化区裸地、草地、灌丛和乔木林4种典型的生境均能广泛分布[5].ZHOU[6]报道了茂兰喀斯特森林区域苔藓植物45科、94属、144种;WU等[7]报道了茂兰洼地内的3科、13属和31种叶附生苔类植物;ZHANG[8]统计了茂兰喀斯特森林生态系统共有28科、93属、186种藓类植物;RAN等[9]探讨了茂兰地区铁皮石斛的生长状况与其伴生苔藓之间的关系;TAN[10]报道了茂兰保护区黄杨沟地区有藓类植物22科、45属、66种、1亚种;LI等[11]在该地区发现濒危物种贵州蓑藓(Macromitrium fortunatii)的新分布;ZHANG等[12]发现了该地区有木灵藓科(Orthotrichales)新种——茂兰蓑藓(Macromitrium maolanense).尽管有关茂兰保护区种子植物的多样性和生态学问题研究广泛[13-17],也有关于苔藓植物区系与分布的初步研究,但是目前缺乏对茂兰地区苔藓植物多样性分布格局及其环境影响因素的科学认识和系统研究.
藓类植物配子体占优势,没有维管组织形成的输导系统,营养主要来自体表吸收,个体小,从而对环境资源要求少,又属于变水植物(poikilohydric plant),具有孢子远距离传播能力,且世代交替短、对基质专一性强、定居和消失速率快,这些特征与维管植物有很大的差异[18].因此,维管植物的众多生态学现象和规律并不一定在藓类植物中呈现[19].研究喀斯特地区藓类植物多样性格局及其形成的环境因素,将为更好地认识藓类植物这一特殊类群的生态分布规律和适应特点,为喀斯特地区藓类植物的多样性保育与利用,特别是在喀斯特石漠化治理中更好地发挥藓类植物的作用提供科学依据.
由于藓类植物对环境变化敏感,在评估景观类型与生境变化对藓类植物群落的影响方面,功能多样性可能具有更好的指示价值.功能多样性主要指某一群落内物种间功能性状的变化范围,或特定生态系统中所有物种功能性状的数值和范围[20-21].有关其测度方法,研究者通常根据实际问题选择较为适合的功能多样性指数,以反映植物功能性状应对不同环境的变化情况[22].MASON等[23]把功能多样性划分为功能丰富度、功能均匀度和功能离散度.有关藓类植物功能多样性的研究,较早的工作是PARDOW等[24]基于苔藓植物生活型研究了热带雨林树附生苔藓植物的功能多样性;BERDUG等[25]在美国云杉林带中,基于形态、生态和生殖有关的多种性状组合,研究了林窗范围冠层梯度上苔藓植物物种和功能多样性.除此之外,很少有关于藓类植物功能多样性的研究.
物种多样性和功能多样性之间存在着一定的关系[26-31],也有学者认为两者在生态系统的功能并不相同,功能多样性的重要性大于物种多样性[32],因此,两者并不是简单的替代关系.在藓类植物类群中开展功能多样性与物种多样性关系的研究,能为更好地认识植物物种与功能多样性的关系提供藓类植物方面的证据.
本文作者主要研究了茂兰保护区藓类植物物种与功能多样性的分布格局,藓类植物物种与功能多样性的关系,以及影响茂兰地区藓类植物物种与功能多样性的主要环境因素,从而为该地区藓类植物多样性保育、发挥藓类植物在喀斯特石漠化治理中的作用提供一定的科学依据.
1 研究区域与方法
1.1 研究区域
茂兰国家级自然保护区位于贵州省南部荔波县境内,地处东经107°52'10″—108°05'40″,北纬25°09'20″—25°20'50″,总面积212.85 km2.海拔430~1 078.6 m,森林覆盖率可达87%.保护区内的成土母岩以白云岩和石灰岩为主构成,岩石裸露率高达80%;土壤层较薄、不连续,其主要为黑色石灰土且较少,多存于石隙石缝处.该地区属于典型的中亚热带季风湿润气候区,年平均气温为18.3℃,全年降水量1 752.5 mm,年平均相对湿度83%,丰富的降水量、适宜的温度和湿度,为各类植物的生长发育提供了有利的气候条件[33].
在对茂兰国家自然保护区地形地貌、主要生态系统和生境类型及分布范围了解和考察的基础上,共选取了石上森林、喀斯特沟谷(青龙潭)、漏斗森林(旺牌山)、洞塘古梅林、村落(尧兰)进行藓类植物系统的调查采样.其中代表性生态系统情况如下:
1)石上森林:属于典型的喀斯特森林地貌,植被总盖度高,几乎达到100%,常为常绿落叶阔叶混交林.林下岩石裸露度达90%以上,并有丰富多样的小生境[34-35].
2)旺牌山漏斗森林:地势起伏大,四周山峰封闭,中部形成深陷陡峻的倒圆锥形洼地,缺水少土,甚至基岩裸露,原生性喀斯特森林保存相对完好,地处保护区核心地段[36].
3)尧兰:喀斯特居民区附近,周缘有耕地,边缘有丘陵小山,该组样地内植物总盖度较低,乔木层不明显,多有裸露的岩石,人为干扰强烈[37].
4)洞塘古梅林:位于原始森林边缘地带的山脚、谷地、台地,生长着年代较久远的野生梅(Armeniaca mumeSibe.et Zucc.),林内石砾多,并伴有裸土、石缝、腐木和古树等,小生境类型较为丰富[38].
5)青龙潭:为典型的喀斯特沟谷,是茂兰地区常见的一种地貌类型,由河流的冲刷侵蚀以及地质运动共同作用形成,该地区小气候相对封闭,水湿条件极好[10].
1.2 野外调查
根据各区域生境类型与分布情况,随机设置样地,样地长宽为2 m×2 m,尽量覆盖不同的生境类型.每个样地范围内的生境尽可能相似.采用Microcoenose法调查每个样地藓类植物盖度.具体方法是:在样地内选取藓类盖度最大的5处设立样方,样方之间的距离不少于0.5 m,以避免重复采样,样方长宽为30 cm×30 cm.调查时,用长宽为33 cm×33 cm的细铁丝筛,筛孔尺寸为3.0 cm×3.0 cm,估测藓类在网格线交叉处出现的次数(每个框架100个交叉点),以此辅助估计每个样方藓类盖度[39].树附生藓类以离地面1.3 m处,胸径大于20 cm的树为采样点,分别在树干离地面30,90,150 cm处,按藓类植物分布较多的一面设立长宽为30 cm×30 cm的样方3个,按上述方法估测样方内藓类植物盖度.同时估测样地环境条件,其中,林冠郁闭度、草本植物盖度用百分制;湿度(由干到湿)、土层厚度(由薄到厚)分别赋值1~5(五级目测),用GPS记录样地经纬度和海拔.
样地内采集的苔藓植物标本鉴定至种,鉴定标准参照中国《中国苔藓志》(第1~8卷)[40-47]、《贵州苔藓植物志》(第一、二卷)[48-49],根据JIA等[50]的中国苔藓植物名录定名.标本存放于上海师范大学苔藓标本室(SHTU).
通过野外与实验室观测,以及查阅文献等方式获取6个藓类植物功能性状指标值(表1).

表1 选取的6个藓类植物功能性状
1.3 统计方法
基于样地中各藓类植物的相对盖度,计算各样地的Shannon多样性指数和Pielou均匀度指数.基于表1中6个功能性状和各样地藓类植物中的相对盖度,分别计算功能丰富度(FRic)、功能均匀度(FEve)、功能趋异度(EDiv)[22-23,51].以样地多样性指标为基础,再计算各区域平均的多样性指标.
各环境因子之间有可能存在多重共线性现象,首先计算了土层厚度、草本盖度、林冠郁闭度、海拔高度、枯枝落叶盖度、湿度这些环境指标间的Pearson相关系数,发现这些指标两两之间的相关系数均小于0.50.因此,分别以这些环境因素单独与多样性指标进行拟合.为在同一坐标图上反映环境因子对藓类植物多样性的影响,对环境变量与多样性指标进行最大值标准化,使其数据处于0~1之间,再分别拟合多样性指标与环境因子之间的关系,探讨影响茂兰苔藓植物多样性的环境因素.
2 结果与分析
在茂兰的石上森林等区域共设立342个样地,其中地面生样地307个,树附生样地35个.从这些样地中共采集1 668份标本,有藓类植物319种,隶属于36科、106属,按种数由多到少的前10个大科分别是 丛 藓 科(Pottiaceae)44种、青 藓 科(Brachytheciaceae)40种、真 藓 科(Bryaceae)28种、蔓 藓 科(Meteoriaceae)23种、灰藓科(Hypnaceae)21种、提灯藓科(Mniaceae)19种、羽藓科(Thuidiaceae)19种、凤尾藓科(Fissidentaceae)19种、绢藓科(Entodontaceae)17种和平藓科(Neckeraceae)15种.
2.1 不同景观的物种与功能多样性
5种代表性景观类型中苔藓植物的物种多样性如图1~2所示.
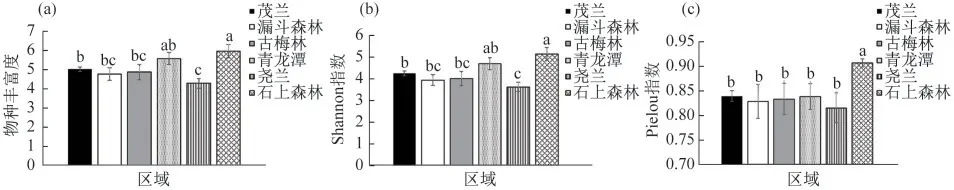
图1 茂兰主要景观的藓类植物物种(a)丰富度、(b)多样性和(c)均匀度(不同字母代表差异显著,下同)
5个景观类型中,青龙潭和石上森林区域的物种丰富度最高,平均每个样点分别为5.58和5.96种,其次为古梅林,为4.87种,再次为漏斗森林和尧兰,分别为4.77和4.29种;Shannon多样性指数以石上森林的最高(5.11),除青龙潭(4.67)之外,明显高于其他几个区域(p<0.05);其次是古梅林(3.98)和漏斗森林(3.92),尧兰最低(3.6),如图1所示;石上森林的物种均匀度(0.91)明显高于其他区域(p<0.05).
功能多样性方面,古梅林的FRic指数最高,青龙潭和古梅林的FEve指数和FDiv指数相对较高,但是这些指标与其他区域的差异不显著(图2).
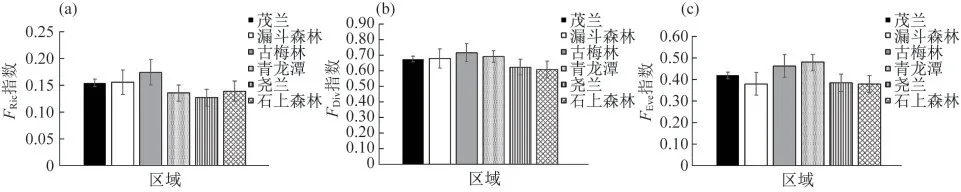
图2 茂兰主要景观的藓类植物的(a)功能丰富度、(b)趋异度和(c)均匀度
2.2 不同基质的藓类植物物种与功能多样性
为反映茂兰藓类植物物种和功能多样性在不同基质条件下的变异特点,分别计算了土生、石生、树生环境下藓类植物的这两个指标.
在茂兰自然保护区,石生藓类植物的Shannon多样性指数最高(4.79),树生藓类次之(4.33),再次为土生(3.12),前两者明显高于后者(图3).石生藓类植物FDiv和FRic指数最高,显著高于树生与土生藓类;就功能均匀度来讲,树生和石生较高,明显地高于土生种类.
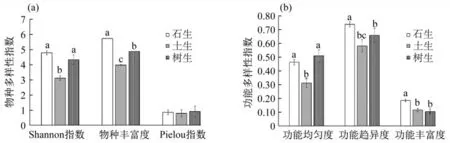
图3 茂兰树生、石生和土生藓类植物的(a)物种多样性与(b)功能多样性
2.3 环境因素对物种与功能多样性的影响
对于地面生藓类,样地林冠郁闭度对其多样性有显著影响.随着郁闭度(x1)的增加,地面藓类植物的物种丰富度(SR)、Shannon多样性(SS)、均匀度(SP)以及功能离散度(FDiv)、均匀度(FEve)均显著上升(p<0.05),但是对功能丰富度(FRci)的影响并不显著(p>0.5)(图4).
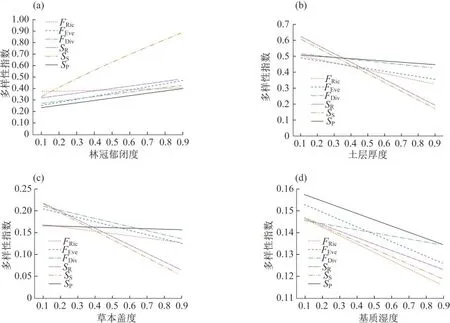
图4 环境因子(a)林冠郁闭度、(b)土层厚度、(c)草本盖度、(d)基质湿度对茂兰藓类植物多样性影响.(数据进行了归一化处理,下同)
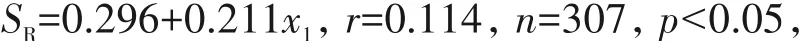
其中,r为拟合优度,用来衡量估计的模型对观测值的拟合程度;n为样本数,即物种数;
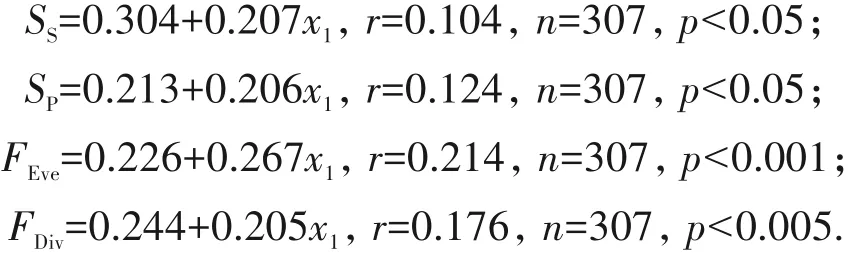
对于地面生藓类植物,随着基质的土层厚度(x2)增加,物种丰富度、多样性,以及功能丰富度、均匀度均呈显著下降趋势(p<0.01),离散度也呈下降趋势(p<0.1),但是物种均匀度下降趋势并不明显(p>0.5)(图4).
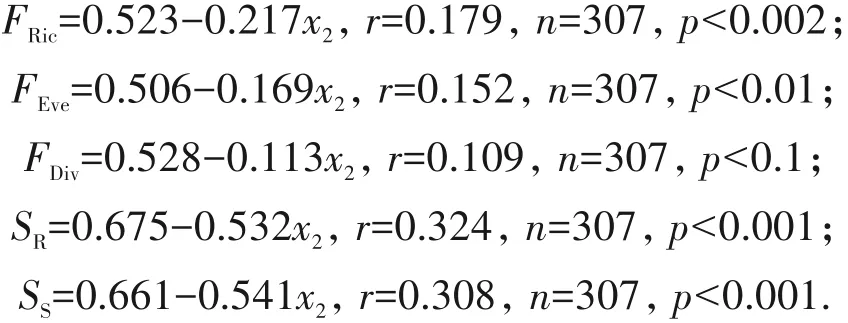
草本盖度对地面藓类植物的多样性也有较大影响.随着草本盖度(x3)增加,地面藓类植物的多样性指标呈不同程度的下降,特别是物种丰富度和多样性、功能趋异度和均匀度,下降显著(图4).
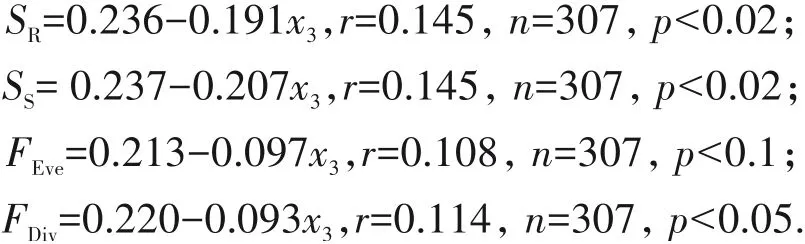
基质湿度对物种丰富度、多样性和均匀度,以及功能趋异度的影响并不显著(p>0.2),但是随着基质湿度(x4)的增加,功能均匀度和功能丰富度显著下降(p<0.05)(图4).
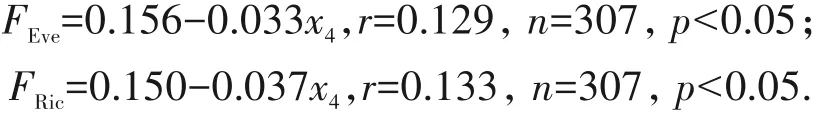
为了解海拔对功能多样性指数的影响,分别对石生、树生和土生基质的藓类植物多样性与海拔的关系进行了拟合,发现石生藓类植物功能丰富度随着海拔的升高而呈现显著增加趋势,而树生藓类的则显著地降低,与土生藓类的关系不明显(图5).
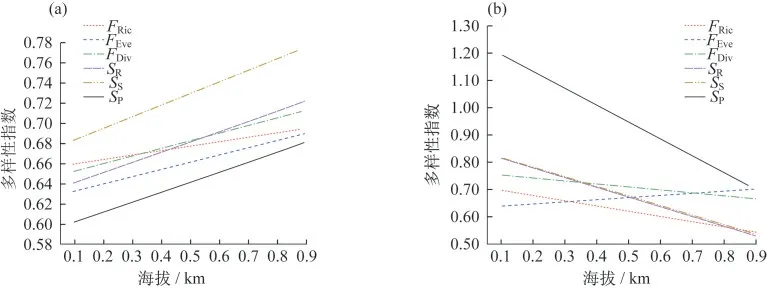
图5 海拔对(a)石生与(b)树生藓类植物多样性的影响
2.4 物种与功能多样性的关系
物种丰富度与功能丰富度、物种多样性与功能趋异度、物种均匀度与功能均匀度之间均呈极显著的正相关关系(p<0.001)(图6).
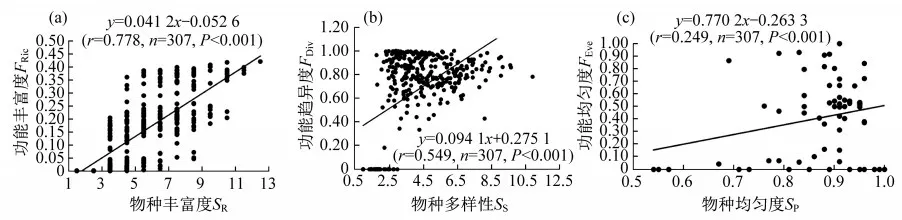
图6 物种多样性与功能多样性之间的相关性.(a)FRic与SR;(b)FDiv与SS;(c)FEve与SP
3 讨论
3.1 不同采样区域的藓类植物多样性
贵州茂兰地区藓类植物物种多样性丰富,其中以石上森林等喀斯特森林地区的藓类植物物种多样性最高.藓类植物的多样性程度与湿度、郁闭度和小生境的多样化程度等显著相关.相对适宜的湿度[52-53]、自然森林周围一定的人为生境[54],和多样化的小生境类型[55-57]适宜更多藓类植物的生长.在石上森林地区,由于其地理位置特殊以及喀斯特地质地貌影响,形成丰富多样的小生境.有岩石裸露、气候变化大的石芽,崩塌的大石块干旱生境;也有土层相对深厚,气候变幅小的石沟、石缝湿润生境.小生境的多样性导致植物群落物种丰富,生态系统结构复杂.同时森林中有一定数量的树附生藓类植物,溪流中又有些水生、湿生藓类植物.石上森林又是重要的旅游景点,构建有步道台阶,为藓类植物的分布提供了不同郁闭度的冠层环境,以及林缘开阔、草本盖度相对较小的生境.结合物种多样性数据,石上森林景观的藓类植物多样性较为丰富,是茂兰藓类植物保护中值得关注的区域.
在茂兰国家自然保护区,不同采样区域间的藓类植物的功能多样性指数虽存在一定差异,但差异不显著.在维管植物类群中,植物功能多样性受地形地貌等区域性因素影响,往往在不同区域间差异显著[24,58-61].茂兰自然保护区不同区域藓类植物的功能多样性没有明显差异,其原因是不同采样区域间海拔、温度等地理和气候因素差距不大.不同的环境条件,例如土地利用方式、人为干扰强度等会影响物种多样性与功能多样性之间的关系.在具有强烈扰动或环境胁迫的环境中,物种的功能属性表现出趋同性,物种性状组成将被限制在适应该扰动或环境选择压力的范围内,导致种间性状差异减小,功能多样性不再增加,也就不能通过物种多样性解释调控生态系统过程的种间功能差异.茂兰地处喀斯特地区,生境的特征表现为干旱缺水,因此性状上有趋同倾向,导致物种多样性区域间差异较大,而功能多样性差异不明显的特征.
3.2 环境因素对藓类植物多样性的影响
藓类植物对基质专一性强,基质类型对藓类植物多样性具有重要的影响.茂兰具有典型的喀斯特地貌,岩石裸露率高达80%,造就石块、石沟、石缝等多种多样的小生境类型,在石漠化严重的区域,因岩溶作用天然形成的千姿百态的石灰岩,在大小、形态、表面粗糙度、土层厚薄、水湿条件、所处环境的郁闭度等方面高度多样,使石生藓类种类多样性、功能多样性、功能趋异度、丰富度和均匀度均相对较高,明显高于土生藓类植物.GUO等[57]在长白山地区的研究得到了相同的结论.茂兰地区具有独特的喀斯特地貌,岩石裸露度高,以及石块、石沟、石缝等小生境多样化程度更高的石生环境,因此石生藓类植物的Shannon多样性指数更高.
随着保护区的设立,茂兰的喀斯特森林得到了有效的保护,在青龙潭沟谷及两侧森林,以及石上森林、漏斗森林等有典型的、发育良好的喀斯特森林.树龄对树干附生苔藓分布影响明显,随着树龄增加,树木胸径增加,苔藓植物定居树干的机会增加.同龄、同一树种、不同植株树干上的附生苔藓植物群落受到了湿度、光照和地形等环境因子的影响而呈现多样化[62].树干位置的高度不同,树皮含水量、周围光照强度、CO2浓度、温度等会有差异,从而分布有不同的苔藓植物种类[59].树皮pH值、缓冲能力、持水性能、树胶、单宁存在与否,以及树皮硬度等对附生苔藓植物的分布也有重要影响[63].在茂兰自然保护区的不同森林内,树的种类多样,树龄与树皮粗糙度各异.尧兰等村落附近古树多,使得树附生藓类植物多样性程度较高.相对于功能丰富度与功能离散度,树附生藓类植物的功能均匀度比石生与土生藓类均要大,可能的原因是,相对于土生和石生环境,树干环境更同质化一些,树附生藓类植物的性状在树干上分布规律性更强些,对树干微环境有效资源的利用更为全面.
就地面藓类植物来讲,随着林冠郁闭度的增加,藓类植物的物种与功能多样性均呈上升趋势.在茂兰,石漠化严重的区域,例如尧兰村落周围的一些区域,林冠郁闭度小,藓类植物也单一,盖度也小;相反,在保护良好的区域的林下石块,例如青龙潭沟谷中的石块,以及这些沟谷两侧的喀斯特森林林下的石砾的郁闭度相对较大,林下石生藓类植物比较丰富.另外,郁闭度越低的地方,往往岩石裸露、光照强烈,环境干旱而严酷.在这样的环境中,藓类植物性状上更趋于耐旱性,不同物种对有效资源的竞争越激烈,所以功能趋异度变低.一般的亚热带常绿阔叶林内,由于郁闭度特别高,林下苔藓植物也并不丰富[64].但是喀斯特森林由于石漠化生境的原因,森林发育并不太好,一般其林冠郁闭度不高,一定范围内,随着郁闭度的增加,形成了适合苔藓植物分布的透光环境.总体上讲,在喀斯特不同程度的石漠化地区,随着石漠化的改善,森林群落的林冠郁闭度增加,藓类植物的物种和功能多样性将上升.随着草本植物盖度,基质中土层厚度、湿度的增加,茂兰喀斯特地区藓类植物的多样性呈下降趋势.可能的原因是,草本植物对苔藓植物具有很强的竞争优势[65],随着样地草本植物盖度的增加,土层厚度也相应的增加,藓类植物表现出物种与功能多样性指数下降的现象.
本次调查有相当一部分样地位于青龙潭沟谷、漏斗森林、石上森林等湿度较大的环境,因此湿度对物种丰富度、多样性、均匀度,以及功能趋异度的影响并不显著(p>0.2).由于部分茂兰地区石生环境众多,在有一定湿度,偏中生化的石生环境中藓类植物种类丰富、微生境更多样化;而一些溪沟环境、湿度特别高的样地,生境相对单一,功能丰富度也低,性状更趋同,功能均匀度也低,因此,藓类植物的功能丰富度和均匀度随着湿度的增加显著下降.
3.3 基质和海拔梯度上的多样性变异式样
石生环境下,随着海拔的升高,草本植物盖度符合二项式函数(r<0.113,n=307,p<0.05),表现出由小变大,再变小的变化式样.在低海拔的区域,不少样地处于喀斯特沟谷,虽然石面草本盖度并不大,但是不少石块处于沟谷溪流中,分布着一些湿生、水生藓类植物,虽然生物量大,但是种类单一,物种和功能多样性均较低;在中等海拔高度的地区,草本植物盖度较大,苔藓植物的多样性也不是很高;到相对高海拔的位置,环境中裸露的石砾较多,草本盖度较小,藓类植物有较多的分布.因此,对于石生基质的藓类植物,其多样性随着海拔的上升而增加.
对于树附生藓类植物来说,随着海拔的升高,森林变小变稀疏,主要的树种变得细而矮小,树附生藓类植物也少.因此,茂兰地区的树附生藓类植物种类随着海拔升高而变少,这与AH-PENG等[66]在热带雨林地区得出的结果相同.
3.4 物种与功能多样性的关系
物种多样性和功能多样性之间存在着一定的关系[26-27],本研究也得出了类似的结果.由于苔藓植物功能性状测量难度大,有学者讨论过是否用便于计算的物种多样性指数来代替功能多样性指数[67-69].但同时,也有学者认为两者对生态系统的功能并不相同,功能多样性的重要性大于物种多样性[28],因此,两者并不是简单的替代关系.但是本研究结果表明,藓类植物的物种多样性与功能离散度、物种丰富度与功能丰富度、物种均匀度与功能均匀度之间均存在极显著的正相关关系.反映出物种多样性可以在一定程度上来表征功能多样性.
4 结论
本文作者以茂兰地区342个样地、319种藓类植物频度、盖度、功能性状数据以及环境因子信息,探究了该地区物种多样性和功能多样性现状、两者关系及影响因素.贵州茂兰石上森林景观的藓类植物多样性较为丰富,是茂兰藓类植物保护中值得关注的区域.不同的基质中,石生和树生藓类的物种多样性高于土生藓类植物;石生藓类植物功能趋异度和丰富度指数最高,树生藓类植物的功能均匀度较高.随着林冠郁闭度的增加,地面藓类的物种多样性与功能多样性均显著上升,但是随着草本盖度、土层厚度和基质湿度的增加,这些指标下降或显著下降.随着海拔的上升,石生藓类植物的物种多样性与功能多样性指标呈现上升趋势,而树附生的呈现下降趋势.茂兰地区藓类植物的物种丰富度与功能丰富度、物种多样性与功能趋异度、物种均匀度与功能均匀度之间均呈极显著的正相关关系.未来苔藓植物相关研究中更应注重探讨环境耦合变化如何改变两者关系,探索相关机制,从而更好地为苔藓植物多样性的保护提供理论支撑.
贵州省是中国苔藓植物多样性最高的省份之一,茂兰国家自然保护区具有典型的喀斯特地貌,通过对茂兰藓类植物标本的采集和鉴定,了解茂兰藓类植物多样性现状,进一步充实贵州省苔藓植物物种多样性,为开展苔藓植物多样性保护提供基础资料支撑.
