北京地区木本春季物候特征与生物学特性的关系
2023-01-13高新月戴君虎陶泽兴
高新月, 戴君虎,3, 陶泽兴
1 中国科学院地理科学与资源研究所,陆地表层格局与模拟重点实验室, 北京 100101 2 中国科学院大学,北京 100049 3 中国-巴基斯坦地球科学研究中心,中国科学院-巴基斯坦高等教育委员会,伊斯兰堡 45320, 巴基斯坦
植物物候直接关系植物个体的生存和繁衍[1—2],进而影响植物群落以及生态系统的结构与功能[3],因此是植物最重要的性状之一[4]。传统植物物候研究多关注物候的年际变化及其对气候因子,如气温、降水、光照等变化的响应特征[5—6]。然而,植物物候发生时间还可能受自身生物学特性,如系统发育和功能性状的影响[7]。
系统发育描述了不同植物的进化关系[8]。系统发育差异可能导致种间的物候差异[8]。根据系统发育保守假说,亲缘关系越近的物种在进化史上的分歧时间越短,因此在性状上保持越相近[8—10]。Davies等基于北美和欧洲地区23个站点4000多个物种的开花始期数据,证实亲缘越近的物种开花时间越相近[10]。Du 等基于《中国植物志》记录的近2万种被子植物的花期差异,也得到相似的结论[11]。然而,对巴罗科罗拉多岛热带雨林的102个树种以及哥斯达黎加布劳利奥卡里约国家公园的35个树种的研究发现,植物开花时间受系统发育的影响较小[10, 12]。Caradonna等比较了北美科罗拉多洛基山地区60个物种的花期数据,也发现系统发育对末花时间和花期长度的影响不明显[13]。这说明植物物候与系统发育的关系在不同地区仍存在较大的不确定性。除此之外,不同物候期对气候变化响应的系统发育保守性也存在较大的区域差异。如植物展叶始期对气温的响应敏感度在哈尔滨地区保守[14],而在青藏高原地区并不保守[15]。
研究植物物候与系统发育关系的方法包括系统发育信号和进化模型[16]。系统发育信号,如Blomberg′s K,是度量不同物种性状在进化树上相似程度的指数。通常而言,K接近1表明性状进化接近布朗运动,具有一定的系统发育保守性。相反,较小的K值说明植物性状的进化速率比布朗运动慢,系统发育可能不保守[17]。但这一度量方法有较大局限性。因为K值较小也可能反映亲缘相近物种的性状总是相对静止,即围绕一个最优目标随机进化,并表现出回归最优值的趋势[16]。这也是植物性状系统发育保守的表现。相比之下,进化模型可区分随机进化过程和进化静止状态[16, 18]。如在Brownian motion(BM)模型中考虑了随机漂变过程(布朗运动),其假设物种间性状的差异程度与其在进化分歧后独立进化的时间成正比[17, 19]。在Ornstein-Uhlenbeck(OU)模型中同时考虑了布朗运动和自然选择力对物种性状进化的共同影响[19—20]。自然选择力的约束作用会使性状向适应环境的最佳状态进化,从而性状可能表现出进化速率比BM过程慢,但保守性强的特征[16, 19, 21]。Glade-Vargas等对南青冈科(Nothofagaceae)27个物种的叶片特征的进化模式进行了分析,发现相对BM模型,OU模型对叶片特征的模拟效果更好[18]。Rafferty等通过文献分析也指出北半球13个植物群落的116个科的植物开花时间的进化更符合OU模型[21]。
植物的功能性状也是另一种重要的生物学特性,它直观反映了植物对环境的适应策略[22]。研究表明,植物的功能性状,如生活型、传粉方式等都可能与植物物候相关[7, 11, 14]。例如,Wang等发现中国新疆干旱区的风媒植物比虫媒植物开花早[23]。对新疆乌鲁木齐地区31种植物的研究发现,乔木比灌木开花早[24]。Panchen等研究发现北半球8个植物园的灌木比乔木展叶早[7]。但也有研究发现,中国亚热带植被区风媒和虫媒植物的开花时间没有明显差异[25]。因此,功能性状与物候的关系仍值得进一步探究。
本研究以北京地区44种木本植物的展叶始期和开花始期为研究对象,分析植物的平均物候期、物候对气温的响应敏感度以及物候发生的积温需求3个物候特征与系统发育和功能性状的关系。首先通过物候特征的系统发育信号和进化模式辨析其是否具有系统发育保守性,然后利用广义估计方程分析物候特征与功能性状的关系。本文的研究成果有助于理解气候增暖背景下物候变化的生物学机制,对物候模型的发展和未来物候格局的预估具有重要意义。
1 数据和方法
1.1 数据来源
北京地区1965—2018年植物展叶始期和开花始期观测数据来自“中国物候观测网(CPON)”。物候观测严格按照《中国物候观测方法》进行[26]。为减少部分观测数据缺失造成的不确定性,在分析前首先剔除观测年份少于15年的物候序列。最终用于分析的木本植物共44种(表1)。与物候期对应年份的北京日均温数据来自中国气象数据网(http://data.cma.gov.cn/)。

表1 研究涉及的物种信息
1.2 研究方法
1.2.1植物物候特征分析
首先,分别计算各物种1985—2018年的多年平均物候期,对季前温度的响应敏感度以及多年平均积温需求。其中,物候期的温度敏感度采用最优时段分析法[27]计算,即以物候期为因变量,对物候影响最大的季前时段的平均温度为自变量的回归方程回归系数。展叶始期和开花始期的积温需求按照如下公式计算。
式中,FGDD为积温需求;t0为积温开始累积日期,设为1月1日;t1表示物候发生日期;x(t)为第t日的平均气温;Tb为积温下限阈值,设为0℃[28]。
1.2.2系统发育树构建
首先,基于The Plant List 网站 (www.theplantlist.org) 查询并核对各物种的学名;然后,利用R程序包V.PhyloMaker中的维管植物巨型骨架树GBOTB.extended.tre构建针对44个物种的系统发育树[29](图1)。

图1 北京地区44种木本植物物候特征在系统发育树上的分布Fig.1 Distribution of phenological traits of 44 woody plants in Beijing on the phylogenetic tree物候特征数值大小与圆圈大小成比例。其中较大的圆圈表示较晚的平均物候期、较大的气温敏感度和积温需求;FLD:展叶始期 First leaf date;FFD:开花始期 First flower date;St:气温敏感度 Temperature sensitivity;HR:积温需求 Heat requirement
1.2.3物候特征的系统发育信号计算
利用Blomberg′s K度量物候特征的系统发育保守性强弱。Blomberg′s K基于布朗运动(BM)模型估算谱系发育信号,该模型预测物种特征分化程度与物种间谱系发育距离成正比[17]。K值大于1表示亲缘相近物种物候特征的相似性比按BM方式进化的相似性更强,相反则更弱,即性状进化可能为随机进化过程或处于相对静止状态。在衡量系统发育信号的显著性时, 将系统发育树上的物种随机交换 1000 次, 分别计算每次交换后的 K 值, 若观察值在a=0.05 的水平上大于零模型的 K 值, 则表明该功能性状有显著的系统发育信号, 反之, 则系统发育信号微弱或没有明显的系统发育信号[17]。具体计算由phytools程序包的phylosignal函数完成。
1.2.4物候特征的进化模式分析
通过对比三种常见进化模型,即BM模型、OU模型和WN模型(假设性状随机进化)对物候特征的拟合效果,探究物候特征最可能的进化模式。模型拟合利用最大似然法,模型效果的检验采用小样本量校正后的赤池信息准则(AICc)。AICc值越小,表明模型的效果越好。具体计算由Geiger包的fitContinuous函数完成。
另外,通过系统发育信号表征曲线(PSR curves)直观表达植物物候特征的进化模式[30]。首先,利用主坐标分析(PCoA)从物种的系统发育距离矩阵中提取若干系统发育特征向量(P1,P2, …Pn),然后依次以特征向量的组合(P1、P1—P2、……、P1—Pn)为自变量,某一物候特征为应变量构建多个系统发育特征向量回归(PVR)模型,绘制PSR曲线。PSR曲线中的x轴为累积特征值,y轴为R2。在布朗运动进化过程假设下(BM模型),PSR曲线呈45°直线型;在随机进化假设下(WN模型),PSR曲线在45°直线下方呈曲线型。若PSR曲线处于两者之间,表明物候特征进化速率比布朗运动慢,但非随机进化,即更符合OU进化过程(OU模型)。基于PSR曲线还可探究植物物候特征进化的平稳性。若PSR曲线在某个点急剧上升,表明突变点对应的几个进化分支(群体)的进化速率差异较大,导致物候特征的种间差异迅速增加[31]。PSR曲线的计算及绘制由PVR包完成。
1.2.5物候特征与功能性状关系研究
本文主要分析2种功能性状——生活型、传粉方式对物候特征的影响。参考《中国植物志》对功能性状的分类如下:①生活型:最大树高超过6米的归为乔木,不超过6米的归为灌木。②传粉方式:裸子植物以及花小且无香味、雄蕊数量较多的被子植物归为风媒植物;花大且有香味、花被片明显、花色鲜艳的被子植物归为虫媒植物[14]。功能性状与物候特征关系的分析采用广义估计方程(GEE)。其原理是将物种间谱系距离矩阵纳入广义线性模型中,并允许在同一模型中包含多个分类和连续型功能性状作为协变量[8, 22]。因此,通过此方法可以去除物种间的系统发育关系的影响,得到功能性状与物候特征之间的真实关系。方程的回归系数描述了不同功能性状类型植物(如乔木和灌木)的物候特征的差异的方向和幅度[22, 32]。具体计算由ape包的compare.gee函数完成。
2 结果与分析
2.1 北京地区植物物候特征
北京地区木本植物的物候期、物候对气温的响应敏感度以及积温需求均呈现较大的种间差异(图2)。展叶始期最早为3月31日,最晚为5月1日。开花始期自3月7日到7月13日,最早和最晚相差超4个月。展叶始期和开花始期的温度敏感度波动范围为-0.2—-8d/℃。就积温需求而言,展叶始期的积温需求处于202—789℃ d之间,开花始期的积温需求相差较大,槐树开花的积温需求(2768℃ d)约为榆树(205℃ d)的13倍。
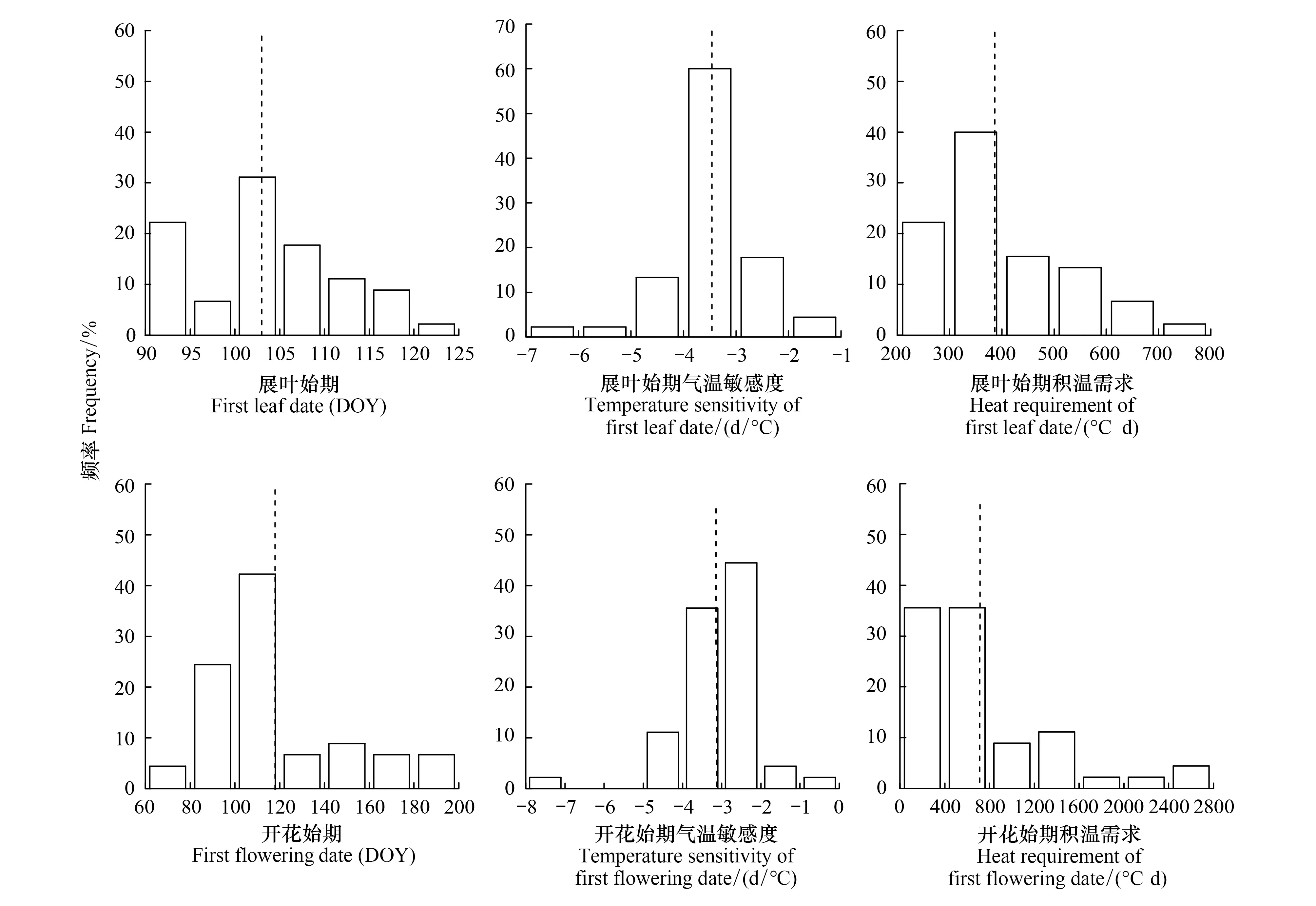
图2 北京地区44种植物3个物候特征的频度分布Fig.2 Frequency distribution of three phenological traits of 44 species in Beijing虚线表示平均值;DOY:序日Day of the year
2.2 物候特征的系统发育信号
植物展叶始期和开花始期系统发育信号显著(P<0.05),说明春季物候期具有系统发育保守性(表2)。其中,开花始期的系统发育信号比展叶始期更强,说明植物生殖生长物候期比营养生长物候期更具有系统发育保守性。相比之下,展叶始期和开花始期温度敏感度的系统发育信号均不显著,说明植物春季物候对温度的响应无系统发育保守性,而两者的积温需求表现出显著的系统发育信号,表明亲缘关系越近的物种积温需求也越相似。

表2 北京地区44种植物物候特征的系统发育信号
2.3 物候特征的进化模式
基于进化模型的拟合优度以及PSR曲线的形状(表3和图3),发现展叶始期、开花始期及其积温需求的进化模式更符合OU模型,表明上述物候特征的进化受到自然选择力的作用,即朝着适应环境的最佳方向进化,这也表示上述物候特征的进化速率比布朗运动更慢,即具有系统发育保守性。就温度敏感度而言,展叶始期的温度敏感度更符合WN的进化模式,说明展叶始期温度敏感度的进化不受系统发育影响。然而,开花始期的温度敏感度更符合OU模型的进化模式,表明开花始期温度敏感度的进化速率比布朗运动假设的进化速率慢,具有较强的系统发育保守性。

表3 北京地区44种植物物候特征的进化模型的AICc值
另外,图3还显示,各物候特征的PSR曲线的上升速率不断变化,说明各物候特征的进化速率具有非平稳性特征。特别值得注意的是,开花始期的温度敏感度的PSR曲线存在若干较为明显的拐点,表明存在几个进化分支(群体),它们的进化速率差异较大,导致物候特征的种间差异迅速增加。
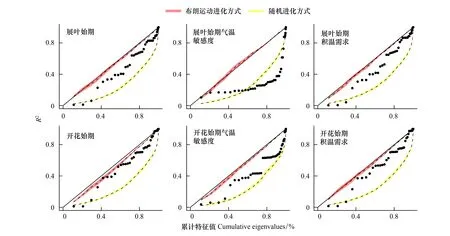
图3 北京地区44种植物物候特征的PSR曲线Fig.3 PSR for phenological traits of 44 species in Beijing红色代表布朗运动进化方式,黄色代表随机进化方式; PSR:系统发育信号表征曲线 phylogenetic signal-representation curves
2.4 物候特征与功能性状的关系
研究发现,植物生活型对展叶始期及其积温需求有显著影响(表3、图4)。其中,灌木展叶的积温需求(平均:362℃ d)比乔木(平均:412℃ d)少约12.1%。灌木的平均展叶始期为4月10日,比乔木早近6天。然而,乔木和灌木展叶始期对气温的响应敏感度(分别为-3.51d/℃、-3.86d/℃)差别不大,说明生活型与展叶始期的气温敏感度无关。植物传粉型与开花始期有显著的相关性。其中,风媒植物的开花始期(平均:4月18日)比虫媒植物(平均:5月4日)早约半个月。但开花始期的气温敏感度和积温需求与传粉型无明显关系。
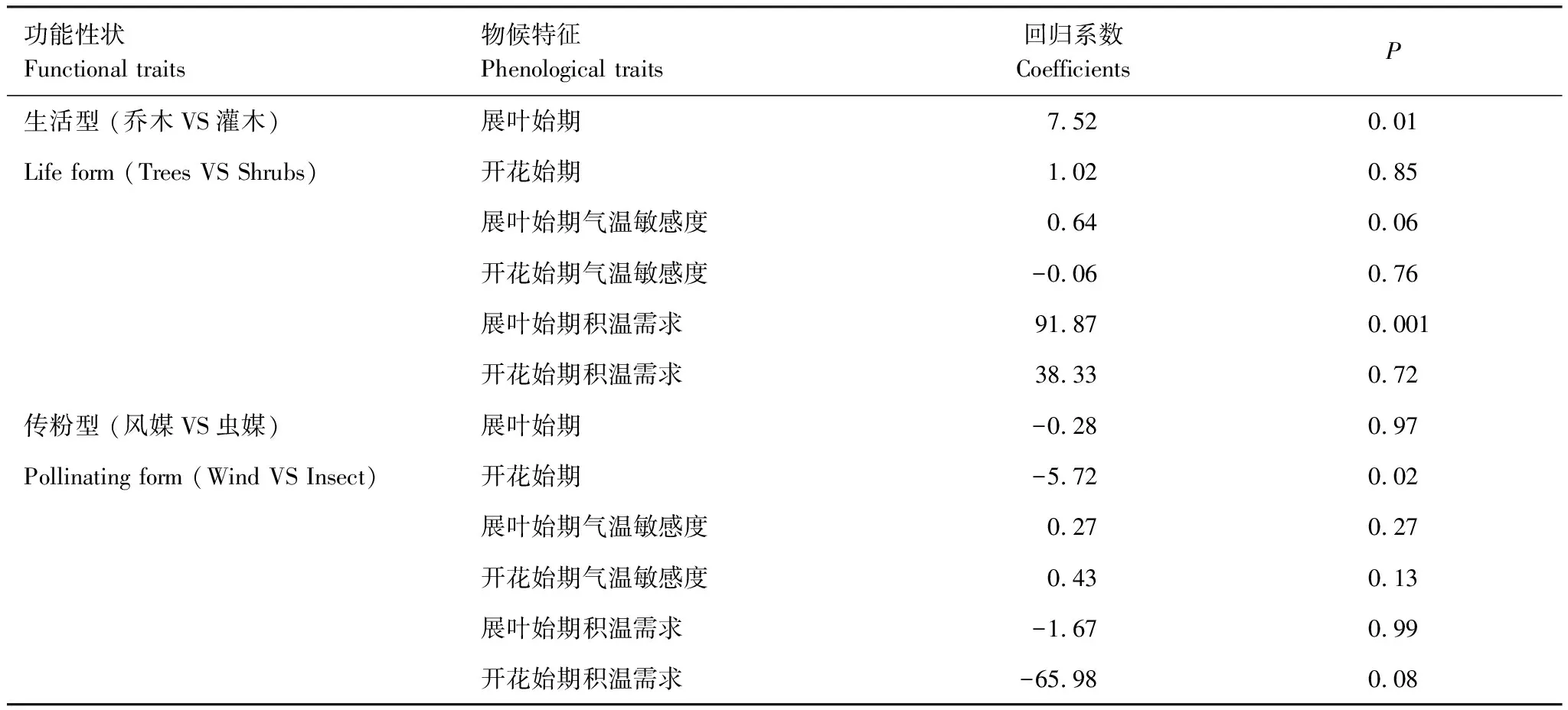
表4 基于广义估计方程的物候特征与功能性状的关系
3 讨论
本文以北京地区44种木本植物的展叶始期和开花始期为对象,研究了系统发育和功能性状与物候特征的关系。与多数研究[10,33—34]结论一致,植物展叶始期、开花始期表现出显著的系统发育信号,表明亲缘关系相近的物种有相似的春季物候期。另外,与Yang 等[15]结论一致,研究也发现春季物候期的积温需求具有系统发育保守性。通常认为,春季物候发生的条件是其积温需求得到满足[35],因此,积温需求的系统发育保守性是春季物候系统发育保守性的解释原因之一。就系统发育强度而言,开花始期及其积温需求的系统发育信号均强于展叶始期,说明开花物候在进化过程中的变异程度较小。可能原因是开花物候直接关系植物个体繁衍[36],若轻易改变可能极大损害植物繁殖能力[13, 37]。而展叶期属于营养生长的一个阶段,间接影响植物个体繁衍[38]。植物通过调节展叶时间响应外界环境条件变化,保证叶片更充分的进行光合作用,并规避遭受的损害风险[38]。
对比三种进化模型对各物候特征的拟合效果,发现OU模型对展叶始期、开花始期及其积温需求进化模式的拟合效果更好,与前人研究发现[18, 21]一致。这表明上述物候特征的进化均受到自然选择的限制,因此进化速率比布朗运动更慢,物候特征具有系统发育保守性。这种自然选择作用可能与植物对环境的适应有关。在长期进化过程中,自然选择作用会“过滤”不适应环境的个体,使群落朝着适应环境的最佳状态去进化[21]。此外,研究表明开花始期温度敏感度的进化模式符合OU模型,即具有系统发育保守性,但系统发育信号不显著。这说明未考虑自然选择力影响的系统发育信号方法可能低估物候特征的保守性。但值得注意的是,PSR曲线显示植物物候特征的进化具有非平稳性(图3),未来仍需进一步完善系统发育保守性的评估方法[16]。
物候特征与功能性状关系的分析结果显示北京地区植物生活型与展叶物候特征显著相关。其中,灌木的展叶时间比乔木早,这与Panchen等[7]在北半球温带地区8个植物园的研究结论一致。可能原因是更早展叶允许灌木在乔木冠层完全形成前充分利用光照进行光合作用[7, 39]。研究还发现植物传粉型与开花始期有显著相关性,风媒植物开花相对更早,这也与Du等对中国19631种植物的研究结论相似[11, 23]。可能的解释有两个方面:首先,风媒树种需要在冠层郁闭前开花,从而减少叶片对花粉随风传播的阻挡[40];其次,北京地区早春相对干冷多风,限制了传粉昆虫的活动,导致虫媒植物开花相对较晚[41]。上述研究结论反映了不同功能性状物种在应对外界环境时的资源投资和收益的权衡策略,同时也在一定程度上体现了植物生态位分化和物种共存的内在机理[22, 24]。
最后,本研究也具有一定的不确定性。受观测记录等限制,现有研究主要选取区域代表性物种进行系统发育分析[24]。这些植物通常跨多科,导致系统发育树的分辨率不够高,在一定程度上影响研究结论的准确性。未来需要加强对北京地区更多树种的物候观测,并尽量细化至某几个科来分析,更深入、准确的探究植物物候特征与生物学特性的关系。
4 结论
本文以北京地区44种木本植物的展叶始期和开花始期为例,探究植物物候与生物学特性的关系,首先,利用系统发育信号Blomberg′s K和进化模型分析了物候性状的系统发育保守性;然后,利用广义估计方程分析了物候特征与功能性状的关系。研究发现:(1)北京地区植物展叶始期、开花始期及其积温需求具有显著的系统发育信号,并且符合OU进化过程,表明上述物候特征的进化受到随机遗传漂变和自然选择力的共同作用,进化速率比布朗运动更慢,具有系统发育保守性。其中,开花始期的系统发育保守性比展叶始期更强。(2)开花始期气温敏感度的进化符合OU模型,说明其具有系统发育保守性;但展叶始期气温敏感度不保守。(3)植物展叶始期及其积温需求与生活型密切相关。灌木的展叶时间比乔木早,积温需求比乔木少。植物开花始期与传粉型显著相关,风媒植物开花比虫媒植物更早。本文的研究成果有助于深入理解物候变化的生物学机制,对丰富物候学理论具有重要意义。
