河流着生藻类多样性对城镇化的响应
——以深圳市为例
2023-01-13刘心怡周佳诚刘振元王伟民陈宇顺
刘 婵, 刘心怡, 周佳诚, 谭 路, 刘振元, 王伟民, 陈宇顺, 唐 涛
1 中国科学院水生生物研究所,淡水生态与生物技术国家重点实验室,武汉 430072 2 中国科学院大学,北京 100049 3 深圳市环境监测中心站,深圳 518049
随着我国经济社会的持续发展,城镇化进程越来越快;据国家统计局2020年公报显示,我国城镇化率已超过60%[1—2]。城镇化为人类生活带来便利的同时,也产生了诸如土地利用类型变更、环境同质化、污染加剧的问题,导致生物多样性降低[3—4]。评估城镇化对生态环境的影响,探讨减缓城镇化对生物多样性影响的有效方法成为日益受关注的问题[5]。
河流与人类关系密切,因此也是最易受人类活动影响的生态系统类型[6]。高度城镇化的河流中,总氮、总磷、溶解性有机质、化学需氧量、电导率等会显著升高,导致水生生物多样性降低,进而影响河流生态系统的功能和服务[7—8]。研究表明,对于不同的多样性指标,城镇化的影响并不完全相同。城镇化对物种丰富度的影响受到研究类群、研究的空间尺度以及城镇化强度的影响[9—10]。例如,多数情况下城镇化会导致鱼类、大型无脊椎动物、藻类等河流生物多样性下降[11—13],但也有研究发现城镇化带来的大量营养物输入会造成细菌多样性和均匀度升高[14]。当然,城镇化引起的环境变化对一些物种带来不利影响的同时也可能有利于另一些物种的生存,因此物种丰富度可能不会出现明显变化。相较于α多样性,β多样性与城镇化强度的相关性更强,城镇化往往会造成群落结构同质化,显著降低β多样性[15—16]。对河流鱼类和大型无脊椎动物的研究发现城镇化不影响物种丰富度,但会引起群落组成和结构的改变[17—18]。有研究发现,城镇化对河流生物α、β多样性的影响与研究区域的环境条件高度相关,存在地区差异[16]。不同地区的河流在气候、地形和地质等自然环境背景的差异导致它们在水化学和水文条件方面存在差异,造成生物群落对于环境胁迫的反应不同[16, 19—20]。因此,城镇化如何影响河流生物多样性尚需更多研究。此外,在水文节律季节性波动明显的区域,季节变化也会对河流生物多样性产生影响,研究城镇化对河流生物多样性影响的季节差异还相对缺乏。
深圳市是我国经济最发达的城市之一,城镇化率已达到100%,土地开发程度达到49%[21],市区河流受人类干扰影响较大,是探讨城镇化对河流生物多样性影响的理想场所。本文利用2020年雨、旱季深圳市主要河流生态调查数据,分析城镇化对着生藻类多样性的影响。着生藻类是河流生态系统的主要初级生产者,对环境变化敏感,常被用作人类干扰指示生物[22]。本研究拟验证以下假说:1、城镇化对不同物种的影响不同,人类干扰使得一些物种消失的同时可能会有新的物种出现,因此城镇化对着生藻类物种丰富度不存在显著影响;但是城镇化造成的水体污染会使一些耐污种成为明显的优势种,使藻类群落的均匀度降低;2、城镇化会造成藻类群落结构同质化,进而降低藻类β多样性;3、旱季河流径流量较小,河流受人类干扰影响更大,因此城镇化对藻类多样性的影响较雨季显著。
1 研究区域
深圳市具有小河众多,大河稀少的特点,流域面积大于1km2的河流有300余条,流域面积大于100km2的河流仅5条,且河道多从市区穿过[23—24]。由于缺乏过境的大江大河作为水源,深圳市河流属雨源性河流,径流量受降水影响较大。深圳市地处亚热带季风气候区,具有明显的旱、雨季,其中4—9月为雨季,12月到来年3月为旱季,其他月份为过渡期。深圳市年平均降水量约为1935.8mm,其中86%集中在雨季[25—26]。深圳市植被覆盖呈现东部大于西部、南部大于北部的地域特征,位于东南部的河流往往具有较高的植被覆盖率[27]。
本研究调查了深圳市九个主要流域:珠江口、茅洲河、深圳湾、观澜河、深圳河、龙岗河、坪山河、大鹏河和大亚湾。其中茅洲河、观澜河、深圳河、龙岗河及坪山河流域面积大于100km2;珠江口、深圳湾和大亚湾水系包括区域内多条相互联通的河流。深圳河、茅洲河、龙岗河、观澜河、深圳湾及珠江口流域位于深圳市中西部,主要流经城区;而坪山河、大鹏河及大亚湾流域位于深圳市东部郊区,植被覆盖率较高。于2020年8月(雨季)、12月(旱季)开展生态调查,于每个流域布设5—9个样点,共计74个样点(图1),两次调查采样点保持一致。根据是否位于城区将74个样点分为城区组和郊区组(城乡结合部样点以建筑、道路等不透水面占比是否超过50%为划分标准),其中城区组包括50个样点,主要位于珠江口、茅洲河、深圳湾、观澜河、深圳河和龙岗河,郊区组包括24个样点,主要位于坪山河、大鹏河及大亚湾流域。两次调查样点因此分成雨季城区组、雨季郊区组、旱季城区组和旱季郊区组。
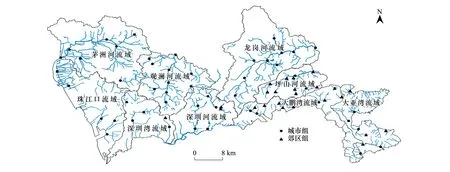
图1 深圳主要城市河流调查样点图Fig.1 Location of the samping sites in Shenzhen main rivers
2 研究方法
2.1 着生藻类采集与鉴定
每个样点调查时,在纵向100m河段内,选取5个不同生境位点,每个位点随机选取3块石头块,用半径2.5cm的塑料圆盖在石块上固定采集区域,用硬毛刷刷取圆盖内区域的着生藻,用水冲洗石块获取藻液。将15块石头上刷取的藻液转移到同一容器中混合均匀,并定容至550mL。现场转移藻液至标本瓶并加入5%甲醛溶液固定保存[28]。着生藻类鉴定在实验室光学显微镜下分两步进行。先在400倍镜下鉴定非硅藻到属并记录硅藻个数,取部分藻液进行酸处理后制作成硅藻封片,参考藻类分类文献[29—36],在1000倍油镜下将硅藻鉴定到种或变种,记录鉴定结果并计算着生藻类相对丰度及物种丰富度(Richness)、香农维纳多样性指数(Shannon-Wiener′s diversity index)、Pielou均匀度指数(Pielou′s index)等α多样性指数。
2.2 环境因子测定
现场采样时,每个点用水质分析仪(YSI)测定水体的酸碱度(pH)、溶解氧(DO)和电导率(Cond),用直读式流速仪测定采样点的流速和水深,并用浊度仪测定水体浊度(Turb)。另外采集350mL水样并加浓硫酸将pH值调至小于2,低温保存并尽快运回实验室用连续流动水质分析仪(SAN++,Skalar)测定总氮(TN)、总磷(TP)、硝态氮(NO3-N+NO2-N)、溶解性无机磷(PO4-P)、高锰酸钾指数(CODMn)、溶解性有机碳(DOC)等水质指标,各指标测定方法参照《水和废水监测分析方法》进行[37]。
2.3 数据分析
分析前根据Pearson相关系数剔除相关性较高的环境因子(r>0.8),并对剩下的环境因子(pH除外)、藻类α多样性指标及相对丰度数据进行log(x+1)转换后进行后续分析。
首先,检验样点位置及季节两因素对环境因子和着生藻类α多样性影响的显著程度。由于各组数据无法同时满足正态分布和方差齐性,使用Scheirer-Ray-Hare非参检验法。该方法是Kruskal-Wallis秩和检验的扩展,主要针对两组以上样本的均值进行统计推断。作为双因素方差分析的非参数替代方法,用于样本无法满足方差分析条件时,检验两个因素的变化是否对样本均值产生显著影响[38]。
其次,使用主坐标分析(PCoA)比较不同样点组间藻类群落结构的差异,并用群落相似性分析(ANOSIM)判断组间差异的显著程度。PCoA分析是一种非约束性数据降维分析方法,用于探究和可视化样本群落组成的差异性或相似性,本研究基于藻类群落的Bray-Curtis距离进行PCoA排序[39]。ANOSIM是一种分析高纬度数据组间相似性的非参数检验方法,它基于样本间距离测度计算关系排名,通过对排名进行置换检验判断样本组间差异是否显著不同于组内差异,常与PCoA等降维分析配合使用来说明样本组间差异的显著性[39—40]。通过PCoA计算得到的物种距离矩阵可以简单地提取质心作为藻类群落的相异性指数,即藻类群落的β多样性[41],绘制质心箱线图并对各组质心进行单因素方差分析以判断四组质心差异是否显著。随后,根据群落组成相异性指数和多点相异性指数,基于导致群落间物种组成差异的两种不同过程,进一步将β多样性分解为物种周转(βsim)和嵌套(βsne)组分,物种周转表示不同群落间的物种替换,而嵌套表示由于物种丧失导致群落间物种丰富度产生差异[42]。并使用置换多元方差分析(PERMANOVA)研究样点位置和季节差异是否对βsim和βsne造成影响。PERMANOVA是根据距离矩阵对总方差进行分解的非参数多元方差分析方法,可以分析不同分组因素对样本差异的解释度,并通过置换检验确定其显著性[40, 43]。
最后,用BIOENV分析和Mantel检验研究环境因子与着生藻类群落结构间的相关性。BIOENV分析通过计算群落结构距离矩阵与环境因子距离矩阵之间的相关系数挑选最适宜的环境因子组合,本研究用Bray-Curtis距离计算群落结构距离,用欧氏距离计算环境因子距离[44]。筛选出重要的环境因子后,使用Mantel检验分析着生藻类群落与环境因子间的关联性,并检验筛选出的各环境因子与城区组、郊区组周转组分的相关性。Mantel检验可分析群落结构差异沿环境梯度的变化程度,Mantel检验中P<0.05说明二者显著相关,而r越大,说明两个矩阵相关性越强[39]。
所有分析均基于R进行。着生藻类α多样性的计算、PCoA分析、ANOSIM分析、PERMANOVA分析、BIOENV分析及Mantel检验使用“vegan”包实现[45]。Scheirer-Ray-Hare检验使用“rcompanion”包实现[46]。β多样性的分解使用“betapart”包实现[47]。
3 结果与分析
3.1 环境状况
Scheirer-Ray-Hare检验结果显示(表1),城区组除pH、溶解氧(DO)和流速(Vel)外的其他环境因子的取值均高于郊区组,其中旱季城区组总氮含量均值高达9.04mg/L,为郊区组(0.92mg/L)的十倍。除总氮外,电导率在城区组和郊区组间也表现出显著差异,无论雨、旱季都存在四倍以上的差异。季节对高锰酸钾指数(CODMn)、DO、浊度(Turb)、水深(WD)和流速有影响。旱季CODMn均值(2.05mg/L)明显高于雨季(1.23mg/L),这种差别在城区组更为明显(3.15mg/L,1.72mg/L)。旱季城区组总氮含量均值(9.04mg/L)高于雨季(6.09mg/L),而郊区组总氮含量无季节差异。两个季节所有样点的总磷、氨态氮、溶解性有机碳、溶解性无机磷和pH平均值无明显区别。此外,比较城区组和郊区组水质指标发现:CODMn和浊度也是在旱季有更大差异。
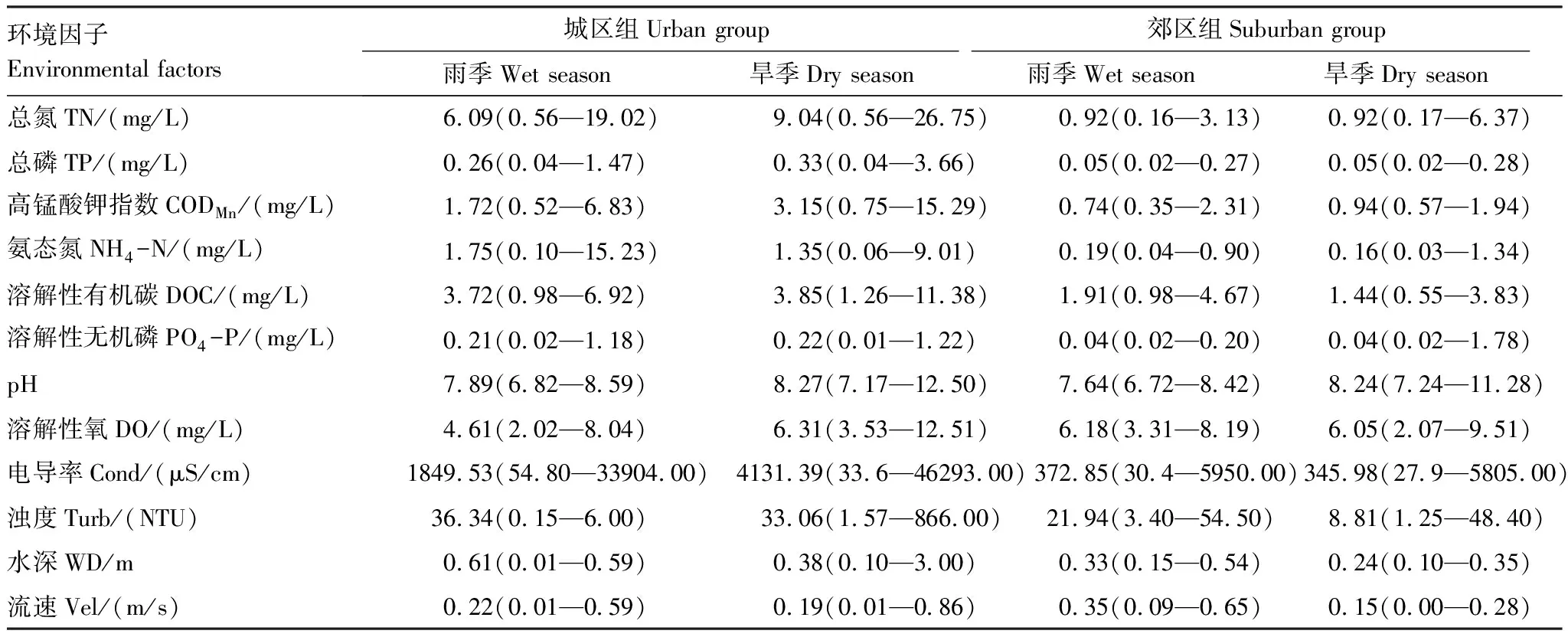
表1 深圳市主要河流环境因子均值及取值范围
3.2 城区与郊区组样点着生藻类α多样性的差别
两次调查共鉴定出着生藻类301个分类单元,分属硅藻门、蓝藻门和绿藻门。其中硅藻门种类最多,为242个分类单元,蓝藻门17个分类单元,绿藻门42个分类单元。雨季鉴定出217个分类单元,其中城区组189个分类单元,郊区组164个分类单元;旱季鉴定出227个分类单元,其中城区组198个分类单元,郊区组161个分类单元。平均相对丰度>5%的物种有9种(表2),其中,颤藻(Oscillatoriasp.)、鞘丝藻(Lyngbyasp.)、席藻(Phormidiumsp.)和微小曲丝藻(Achnanthidiumminutissimum)在两个季节的城区组和郊区组都是优势种,而罕见曲丝藻(Achnanthidiumexiguum)只在城区组作为优势种出现,比利牛斯山曲丝藻(Achnanthidiumpyrenaicum)只在旱季郊区组为优势种,两栖菱形藻(Nitzschiaamphibia)和平庸菱形藻(Nitzschiainconspicua)只在旱季城区组为优势种。比较雨、旱两季优势种相对丰度发现,雨季丝状蓝藻相对丰度较旱季高;综合两季数据来看,城区组相对丰度较高的硅藻门物种主要是菱形藻属,而郊区组相对丰度较高的硅藻门物种主要是曲丝藻属。平均相对丰度大于1%的物种有17种,其中硅藻门12种,蓝藻门3种,绿藻门2种。其他284种藻类的平均丰度小于1%。雨季城区组的指示物种有β-中污带指示种变异直链藻(MelosiravariansC.Agardh)、钝脆杆藻(FragilariacapucinaDesmazières)、克罗顿脆杆藻(FragilariacrotonensisKitton)、尖针杆藻(UlnariaacusKützing)、隐头舟形藻(NaviculacryptocephalaKützing)和椭圆波缘藻(CymatopleuraellipticaW.Smith),以及梅尼小环藻(CyclotellameneghinianaKützing)、平庸菱形藻等富营养化指示种;与雨季城市组相比,β-中污带指示种椭圆波缘藻不在雨季郊区组出现,且大部分污染指示种相对丰度有所下降;旱季城区组的指示物种有β-中污带指示种钝脆杆藻和隐头舟形藻,及富营养化指示种梅尼小环藻、平庸菱形藻和库津细齿藻(DenticulakuetzingiiGrunow);旱季郊区组指示物种有β-中污带指示种隐头舟形藻,以及富营养化指示种梅尼小环藻、平庸菱形藻。α-中污带指示种谷皮菱形藻(Nitzschiapalea)和颤藻、清洁指示种微小曲丝藻在四组样点都存在,其中谷皮菱形藻在城市组相对丰度更高,颤藻在雨季组相对丰度更高,而清洁种微小曲丝藻在郊区组有更高的相对丰度。比较各组出现的污染和清洁指示种的数量和相对丰度,可以发现污染指示种更倾向于在雨季、城市组出现,而清洁指示种更倾向旱季、郊区的环境。
所有样点的着生藻类平均分类单元数为31(取值范围为10—53)(表3),香农维纳多样性指数平均值为2.87(1.54—3.49),Pielou均匀度指数平均值为0.84(0.49—0.94)。三项α多样性指数在城区、郊区组间及季节间差异均不显著。
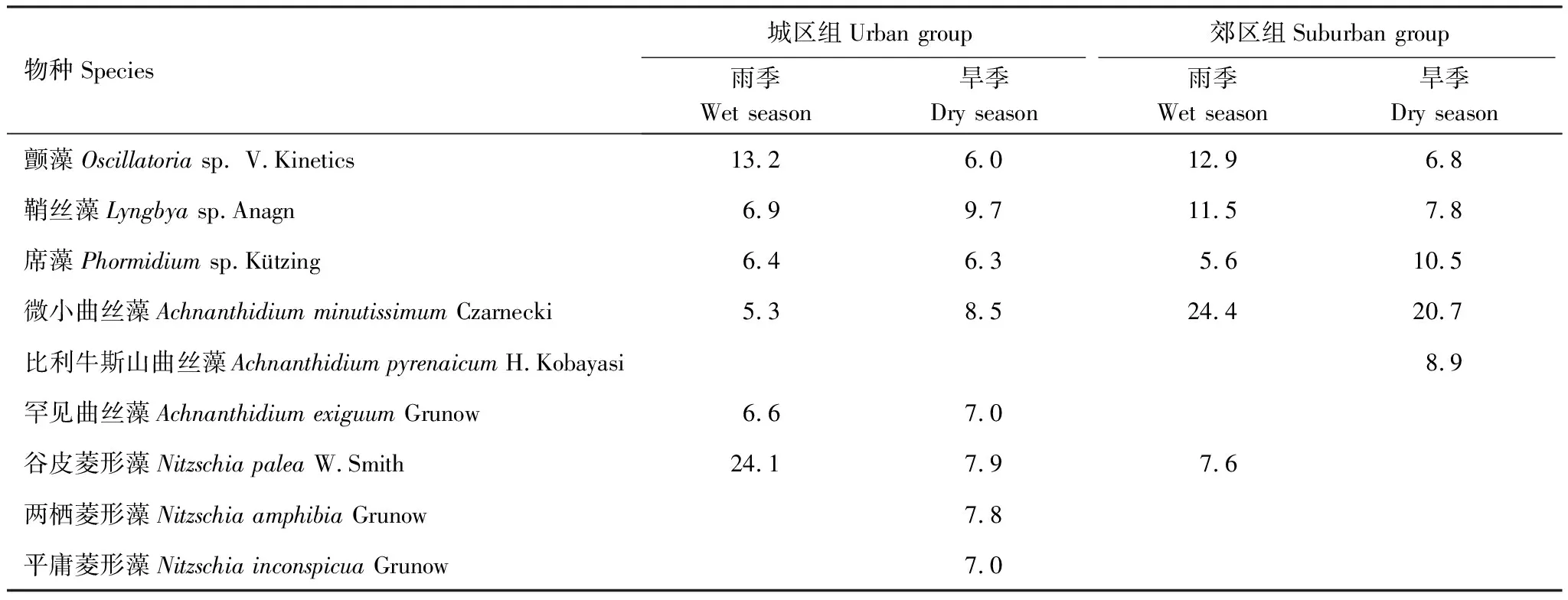
表2 深圳主要河流城区组和郊区组着生藻类优势种相对丰度/%
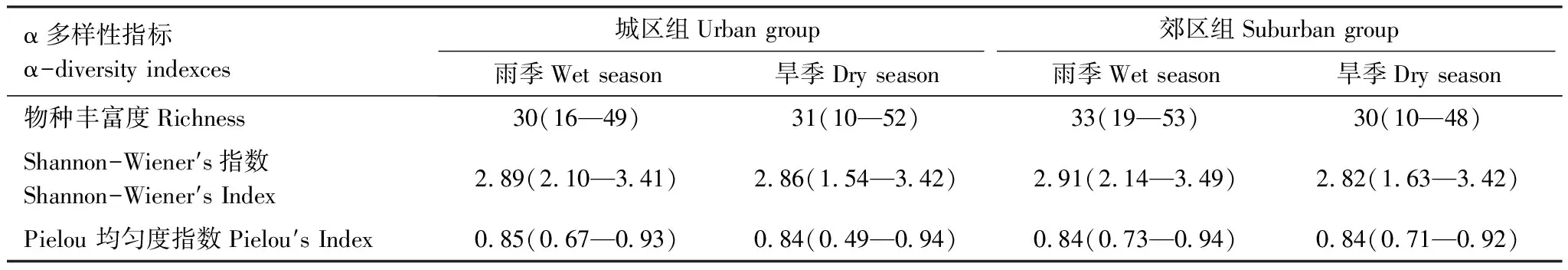
表3 各组α多样性平均值及取值范围
3.3 城区、郊区组样点着生藻类群落结构的差别

图2 基于藻类群落Bray-Curtis距离的主坐标分析(PCoA)排序图Fig.2 Principal Coordinate Analysis (PCoA) plot based on Bray-Curtis distances of algae communities
四组样点在PCoA排序图上分散分布(图2),其中相同季节的城区组、郊区组样点着生藻类群落在PCoA轴1上分离,相同位置不同季节的着生藻类群落在PCoA轴2上分离。ANOSIM结果进一步显示,着生藻类群落结构组间差异显著大于组内差异,分组因素显著影响藻类群落结构(r=0.345,P=0.001)。
PCoA质心箱线图显示(图3),雨季藻类群落β多样性较低,旱季较高;其中,雨季城区组和郊区组β多样性差别不大,旱季郊区组β多样性比城区组高。无论是雨季还是旱季,郊区组藻类群落β多样性的中位数都较城区组高。四组质心方差分析结果显示:雨季城区组质心显著低于旱季城区组(P=0.015)和旱季郊区组(P=0.022),其他各组间质心差异均不显著。

图3 藻类群落PCoA质心箱线图Fig.3 Boxplot of centroids of PCoA of algae communities
对着生藻类群落的β多样性进行分解发现其空间周转组分远高于嵌套组分(βSIM=0.9745,βSNE=0.0057)。PERMANOVA结果显示城镇化与季节都对βsim产生显著影响(城镇化:R2=0.1058,P<0.001;季节:R2=0.1081,P<0.001),而不同分组间的βsne差异不显著。
3.4 城区、郊区组着生藻类群落与环境因子关系差异
用BIOENV筛选出与各组藻类群落结构显著相关的环境因子(表4)。其中,城区组雨季为TN、CODMn、Cond、Turb和水深,旱季为TN、CODMn、Cond和pH;郊区组雨季为PO4-P,旱季为TN和Cond。对筛选出的环境因子组合进行Mantel检验,结果显示(表4):筛选出的环境因子组合对于雨季组的解释率均低于旱季组,对于城区组的解释率高于郊区组。

表4 藻类群落与环境因子相关性Mantel分析
将筛选出的环境因子分别与城区组、郊区组周转组分进行Mantel检验,结果显示城区组、郊区组周转组分分别与筛选出的城区组、郊区组环境因子显著相关,其中对城区组周转组分解释率最高的环境因子是Cond(P<0.001,r=0.524),对郊区组周转组分解释率最高的环境因子是PO4-P(P=0.01,r=0.169),总体来看环境因子对城区组周转组分的解释率高于郊区组周转组分。
4 讨论
本研究发现,流经城区的河流TN、TP、CODMn、Cond、Turb等指标均显著增高,可见人类干扰对河流水质有显著影响;季节因素对CODMn、DO、Turb、水深和流速产生影响,其中CODMn和Turb均是旱季明显高于雨季,而DO、水深和流速存在季节差异的同时也受到样点位置的影响。深圳市河流是雨源性河流,与雨季相比,旱季径流量更多来自生产与生活污水,因此旱季CODMn和Turb更高也与城市人类活动密切相关。DO在雨季和旱季时具有不同的差异特征,雨季时DO城区组显著低于郊区组,而旱季时郊区组具有较低的DO,这可能是因为雨季时城区组水深较深而流速较低引起的水气交换速率降低导致水中DO浓度降低,且雨季水体中充足的营养物质使得浮游藻类呼吸作用显著增加,也会大量消耗DO[48—49]。郊区组样点多位于河流源头,城区组样点多位于河流中下游,因此城区组具有较大水深,雨季的大量降水使得差距更为显著。对于流速来说,郊区样点间落差较为明显,在水源充沛的雨季具有较高流速;而旱季时由于河流源头处流量很小,郊区组流速也低于城区组样点。从着生藻类群落组成上看,城区组优势种以菱形藻属为主,郊区组以曲丝藻属为主,城区组污染指示种种类数和相对丰度都比郊区组更高。大量研究表明,菱形藻属与水体营养状态相关,其中城区组相对丰度最高的硅藻门优势种谷皮菱形藻对高浓度营养盐污染具有较强的耐受性[50—51],而郊区组主要优势种极小曲丝藻更偏爱低营养盐的生境[52]。城区组显著增高的TN、TP等营养盐指标也从侧面印证城镇化显著影响河流水质,从而对着生藻类群落组成产生影响。对着生藻类α多样性的比较研究发现,样点位置和季节都对藻类α多样性无明显影响,这与本研究的假设不完全相符,可能与城区组和郊区组之间较大的样点数差异有关。
PCoA质心箱线图显示,与郊区组相比,城区组着生藻类群落β多样性较低,这种差异在降水量少、径流主要来源于生产生活污水的旱季更加明显。BIOENV与Mantel检验结果显示:影响城区组着生藻类群落的环境因子主要是TN、CODMn、Cond、pH、Turb和水深,影响郊区组着生藻类群落的环境因子主要有TN、Cond和PO4-P,这与前人研究结果相似[53]。结合城镇化对环境因子影响的分析结果,说明城市人类活动导致的TN、CODMn和Cond水平的显著升高会导致着生藻类β多样性下降,这与其他类群研究中得到的城镇化会造成物种同质化的结论一致[54]。城镇化对着生藻类群落的影响在旱季更加明显可能是因为旱季河流径流缺少雨水汇入,受人类活动干扰更大,环境因子受到的影响更加显著。与旱季相比,雨季藻类β多样性较低,但季节引起的差异比样点位置引起的差异更小。雨季藻类β多样性较低可能是雨季水文条件变化对着生藻类生长的影响导致的,雨季城区组中Turb和水深对着生藻类群落结构具有显著影响的结果也印证了这一点。雨季时,大量雨水汇入河流使得Turb和水深显著增大,水体Turb较大时会降低水体透明度,对着生藻类的生长产生不利影响,水深对着生藻类的影响也与Turb类似,同时水流冲刷也使得藻类β多样性降低。对着生藻类群落与环境因子关系的研究还发现,影响城区组与郊区组的环境因子有所不同,而雨、旱季影响藻类群落组成的环境因子差异较小,说明城镇化对环境因子与着生藻类群落的关系的影响比季节因素更显著。比较影响城区组和郊区组着生藻类群落的环境因子可以发现,TN和Cond对城区组和郊区组藻类群落均有影响,且两个因子取值水平在城区组和郊区组差异显著,是造成城区、郊区着生藻类群落结构差异的主要环境因子;同样结合雨季组、旱季组环境因子差异和着生藻类群落分析结果可以发现造成雨季组、旱季组着生藻类群落结构差异的主要环境因子为CODMn。对β多样性进行分解后发现组间着生藻类群落结构差异的主要来源为物种周转或替换,这与前人研究结果相似,而高于平均水平的βsim说明环境过滤、竞争和扩散限制是形成深圳市河流β多样性的主要机制[55—56]。随后对βsim进行PERMANOVA分析的结果说明城镇化和季节因素共同导致组间β多样性差异,这与前文得到的结果一致。
本研究发现城镇化造成河流TN、PO4-P、CODMn、Cond、Turb等指标水平显著增高,导致河流着生藻类群落β多样性降低,说明城市人类活动干扰会对河流生物多样性带来不利影响。为有效保护河流生物多样性,有必要从源头控制工业和生活污水的排放,减少对河流的污染。尤其是在降雨量较小的旱季,更需要采取措施避免污水排放进入河流,并尽量减少雨水进入河流时带入面源污染。
