普通白菜CYP79B2同源基因的克隆与表达
2023-01-05胡琪侯玉翔李璿李梅兰
胡琪 侯玉翔 李璿 李梅兰
(山西农业大学园艺学院,晋中 030800)
普通白菜(Brassica rapassp.chinensisMakino)又称青菜、油菜,属十字花科芸薹属芸薹种,含较多蛋白质、维生素等成分,营养价值较高,在全国南北各地均广泛栽培[1]。白菜是种子春化型蔬菜,春季容易先期抽薹,因此,研究植物的开花机制具有重要的实践价值。
植物开花是由内源激素和外部环境信号共同作用的结果,已知的五大类生长激素包括赤霉素、生长素、脱落酸、乙烯和细胞分裂素,均与春化作用和成花诱导有关[2],其中生长素(IAA)属于生长促进剂,对植物的生长发育具有调控作用[3]。植物体内IAA 的合成非常复杂[4],其生物合成途径主要分为依赖色氨酸和非依赖色氨酸2 条途径。根据合成的中间产物,依赖色氨酸途径又分为4 条支路,吲哚乙醛肟途径[5]、色胺途径[6]、吲哚乙酰胺途径[7]和吲哚丙酮酸途径[8]。吲哚乙醛肟途径是芸薹属植物中最重要的一条途径[9],又称为CYP79B 途径。CYP79B2 可以催化色氨酸转化形成吲哚乙醛肟,而吲哚乙醛肟是IAA 和吲哚族硫苷的共同前体物[10-11]。Mikkelsen 等[12]发现经1 μmol/L 2,4-D 处理的拟南芥CYP79B2被诱导表达,导致吲哚硫苷积累量增加。杨子琴等[13]发现阻断龙眼顶芽IAA 的运输可导致延迟开花,而IAA 浓度较低时可促进开花。Hull 等[14]发现拟南芥CYP79B2可以将色氨酸转化为吲哚-3-乙醛肟,且过表达CYP79B2拟南芥产生抵抗毒性的色氨酸类似物,表明CYP79B2可能与植物的防御机制和逆境胁迫有关。另有研究表明,CYP79B2在不同植物生长发育各阶段的表达模式既有相似性又有特异性,在菘蓝和大白菜中,经茉莉酸甲酯胁迫,CYP79B2表达增强,但经水杨酸胁迫后,其表达趋势相反[15-16]。王梦雨[17]还发现BoaCYP79B2和BoaCYP79B3的表达量与芥子油苷的含量密切相关,表明CYP79B2在植物体内具有抗癌防癌的作用。
CYP 酶是一类膜蛋白超家族酶系[18]。目前,在拟南芥、油菜、西兰花等植物中已经克隆到CYP79家族中的多个基因,如CYP79A、CYP79B、CYP79D和CYP79F等[19-22]。前期研究主要阐明了CYP79 在植物生长素合成及其对害虫、病原菌的防御机制中发挥重要的作用。除此之外,宋延宇等[23]发现调控薹菜CYP79B5的表达能改善蔬菜的风味品质。何荟如等[24]发现茶树CYP79A1的表达量与品种和摇青程度有关。由此可见,CYP 酶几乎是“万能的生物催化剂”。但前人的研究多集中在生物合成关键酶的表达调控,如硫苷[25]的合成,以及参与多种乙醛肟类化合物的反应调控,如吲哚乙醛肟[26]的转化。而有关CYP79B2在普通白菜开花调控中的研究却鲜有报道。
本研究以普通白菜品系‘75#’为试验材料,通过RT-PCR 方法克隆普通白菜中生长素CYP79B2同源基因,利用生物信息学方法对其序列及蛋白结构进行分析,运用RT-qPCR 方法分析该基因在不同组织和不同时期的表达情况,以期为后续普通白菜开花调控的基因功能验证提供理论依据。
1 材料与方法
1.1 材料
供试材料为普通白菜品系‘75#’,在适宜环境下,种子露白后,4℃低温处理20 d 即可完成春化,然后移栽至穴盘,并分别对移栽后0、10、15 和16 d 的植物组织茎尖进行取样,样品分别标记为V0、V10、V15 和V16。取样量为0.1 g,液氮速冻后于-80℃保存备用。
1.2 方法
1.2.1BrcCYP79B2-2的克隆
1.2.1.1 总RNA 的提取和cDNA 的合成 将V10 样品在液氮中迅速研磨成粉末,按照RNAprep Pure Plant Kit(TIANGEN,DP432)试剂盒提取RNA,使用PrimeScriptTMRT reagent Kit(Perfect Real Time)(TaKaRa,RR037Q)试剂盒将其反转录成cDNA。
1.2.1.2 基因克隆和序列分析 使用大白菜数据库(http://brassicadb.org)查找CYP79B的同源序列,用Primer 5.0 在线软件对基因设计引物(F:5′-ATGTATCTCCTCACAACGCTCCA-3′;R:5′-TTACTTCACCGTCGGATAAAGTTGC-3′)。以1.2.1.1 反转录的cDNA 为模板,使用TaKaRa LA Taq®(TaKaRa:RR02MQ)试剂进行PCR 扩增,其反应体系为Ta-KaRa LA Taq 0.5 μL、LA Taq Buffer II(10×)5 μL、正、反引物以及cDNA 模板各1 μL、无菌水33.5 μL。反应程序为94℃ 5 min;94℃ 30 s,56℃ 30 s,72℃ 2 min,共35 个循环;72℃ 5 min,4℃保存。用1%琼脂糖凝胶电泳检测,条带大小正确且无杂带即可进行回收。
将回收的DNA 产物与pUCm-T Vector(PCR Products Cloning Kit Sangon Biotech,B522213)在PCR仪中连接。后转化至大肠杆菌E.coliDH5α Competent Cells 中,挑选阳性克隆进行PCR 验证,送生工生物工程(上海)股份有限公司进行测序。
1.2.2 生物信息学分析 使用DNAMAN 软件将测序序列翻译成氨基酸,以NCBI 数据库为基础,进行氨基酸序列多重比对,并构建系统进化树。采用在线工具ProtParam 分析氨基酸的理化性质,再分别使用SOPMA 和Swissmodel 在线软件对蛋白质结构进行有效预测,了解蛋白质的一些功能。
1.2.3 基因的表达分析 为更清楚地了解CYP79B同源基因的功能,对普通白菜‘75#’的根、茎、叶、花蕾和果荚进行取样,用RT-qPCR 方法研究其在不同组织中的表达情况。设计BrcCYP79B2-2的特异性引物序列(F:5′-TTAGGTACGGCGTTGACCAC-3′和R:5′-TCACCGACCATAACCAACGG-3′)。
2 结果
2.1 基因克隆
以cDNA 为模板,进行扩增,获得一条与目的条带大小一致的条带(图1)。经测序验证后,基因长度为1 554 bp,命名普通白菜CYP79B的同源基因为BrcCYP79B2-2。提交序列至GenBank,登录号为OM339440。

图1 BrcCYP79B2-2 的扩增电泳图Fig.1 Amplified electrophoresis of BrcCYP79B2-2
2.2 序列分析及多重比对
使用DNAMAN 软件对BrcCYP79B2-2的氨基酸序列进行分析,结果表明,BrcCYP79B2-2共编码517 个氨基酸。多重序列比对结果(图2)表明,BrcCYP79B2-2编码氨基酸序列同大白菜(Brassica pekinensis)、甘蓝型油菜(Brassica napus)、甘蓝(Brassica oleraceavar.capitata)、芜菁(Brassicarapa)、花椰菜(Brassica oleraceavar.botrytis)、萝卜(Raphanus sativus)和拟南芥(Arabidopsis thaliana)具有较高同源性,同源性分别为99.42%、99.23%、98.84%、99.42%、98.84%、97.10%和93.44%。表明克隆基因确实为普通白菜BrcCYP79B2-2,且该基因在不同变种白菜中,序列高度保守。

图2 不同物种CYP79B2-2 氨基酸序列的多重比对Fig.2 Multiple alignment of CYP79B2-2 amino acid sequences from different species
2.3 蛋白质亲缘关系分析
为进一步了解BrcCYP79B2-2 在不同物种之间的亲缘关系,使用DNAMAN 软件构建系统进化树,与大白菜(Brassica pekinensis)同源性最高,并且同花椰菜(Brassica oleraceavar.botrytis)和甘蓝(Brassica oleraceavar.capitata)同属于一个小分支,与萝卜(Raphanus sativus)、白芥(Sinapis alba)和拟南芥(Arabidopsis thaliana)亲缘关系较近,与番木瓜(Carica papaya)、黄麻(Corchorus capsularis)、茶花(Camellia sinensis)等植物的亲缘关系较远(图3)。

图3 不同物种CYP79B2-2 同源蛋白的系统进化树Fig.3 Phylogenetic tree of CYP79B2-2 homologous proteins from different species
2.4 蛋白质理化性质分析和结构预测
BrcCYP79B2-2编码517 个氨基酸,分子量为58.522 29 kD,其理论等电点为8.71,脂肪指数为88.47,亲水性的平均值为-0.167;带负电荷的残基总数为58,带正电荷的残基总数为64,不稳定性指数(II)为36.99,此蛋白质分类相对稳定。
使用SOPMA 和Swiss model 分别预测蛋白质的二、三级结构,结果(图4)显示,BrcCYP79B2-2二级结构有α-螺旋、β-折叠、β-转角和无规则卷曲,主要二级结构是α-螺旋(47%)和无规则卷曲(36.36%);其三级结构模型与二级结构分析结果吻合。
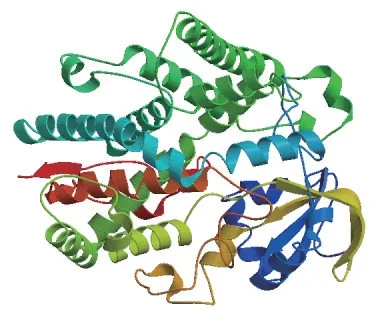
图4 BrcCYP79B2-2 蛋白质三级结构预测Fig.4 Protein tertiary structure prediction of BrcCYP-79B2-2
2.5 基因表达分析
为了解BrcCYP79B2-2在普通白菜不同组织与不同生育期中的表达模式,分别对根、茎、花蕾和果荚组织部位取材,使用RT-qPCR 方法对BrcCYP79B2-2的表达进行定量分析。结果(图5-A)显示,BrcCYP79B2-2在不同的组织中均有表达,在植物茎中的表达量最低,在花中的表达量较低,而在根中的表达量最高。比较茎尖在不同发育期的表达,发现经低温处理后,BrcCYP79B2-2表达较低,在移栽10 d 后达到顶峰,此时为营养生长期,移栽15 和16 d 后,即花芽即将分化和分化开始时处于相对较低的表达水平,说明该基因对低温处理有响应,在营养生长时期茎尖的表达较高,但是在茎尖分化时下降(图5-B)。
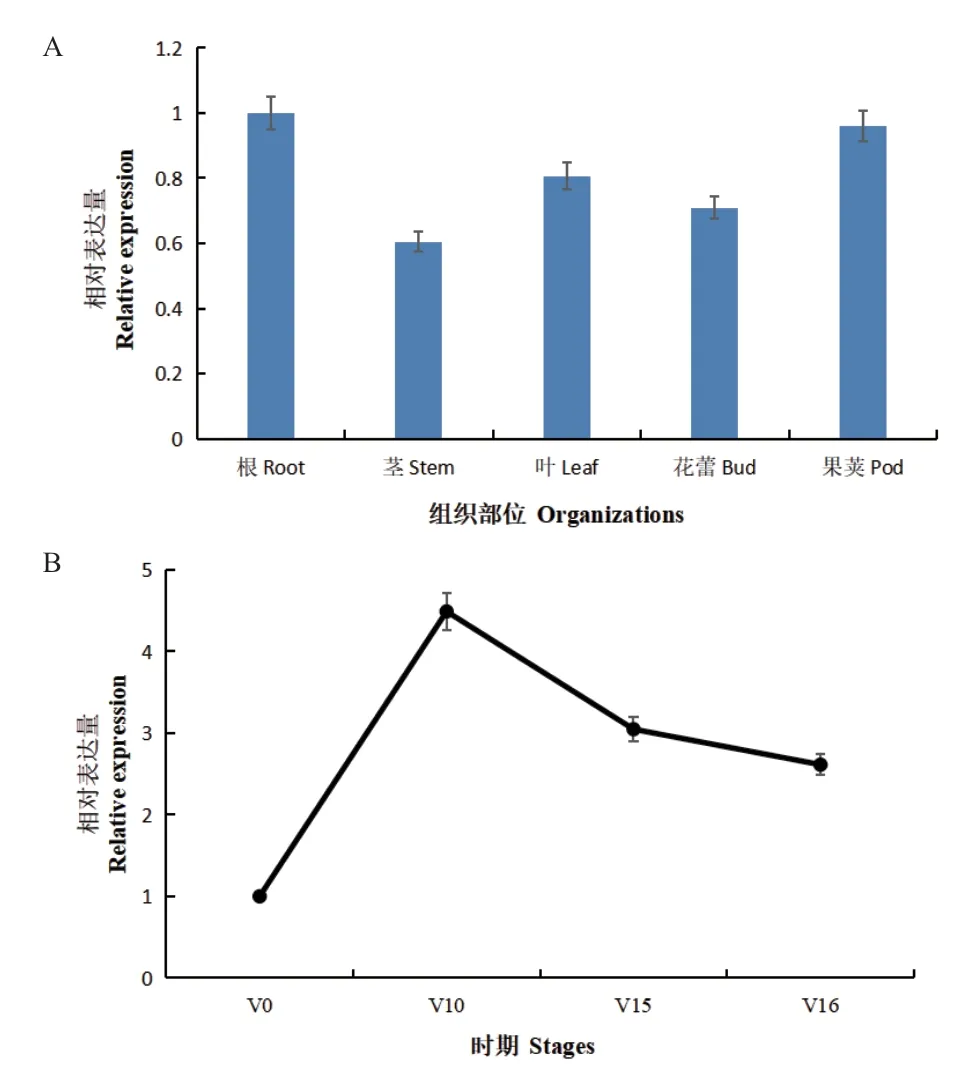
图5 BrcCYP79B2-2 在不同组织的表达和低温处理后不同时期茎尖部位的表达Fig.5 Expressions of BrcCYP79B2-2 in different tissues and in stem tip in different stages after low temperature treatment
3 讨论
本试验利用同源克隆技术成功克隆到普通白菜BrcCYP79B2-2,从蛋白质亲缘关系图来看,普通白菜BrcCYP79B2同源基因所编码的蛋白质与大白菜、拟南芥和萝卜等十字花科亲缘关系较近,而与黄麻、茶花等植物的亲缘关系较远,进一步说明BrcCYP家族基因具有高度保守性。
植物在适宜的外部环境中,生长到一定阶段的时候,其形态结构和生理结构均会发生改变,其中由营养生长到生殖生长的转变过程最为典型[27-28],这个过程即高等植物的开花过程。白菜在开花过程中,激素调节的作用不可忽视[29]。IAA 是五大激素之一,几乎调控植物生长发育的所有方面,包括形态发生和对环境变化的响应[30]。在拟南芥中CYP79B共鉴别出2 个基因:CYP79B2和CYP79B3,它们都能催化IAA 的合成,可能与植物开花有关。CYP79B2过表达是导致植物体内IAA 升高的原因之一[31]。当它们被敲除后,cyp79b2cyp79b3双突变体则表现出叶片短小,体内生长素水平下降的明显特征[32]。本试验通过RT-qPCR 研究在普通白菜不同器官中的表达情况,结果显示,BrcCYP79B2-2在根中的表达量最高,这与孙夏莉等[33]研究结果一致。BrcCYP79B2-2在不同组织和不同发育期的表达含量不同,同样,原晓龙等[34]研究显示蒜头果实中CYP79的表达量在花谢后1月>2月>4月>3月,也进一步说明生长素在植物体内的表达具有时空特异性。杜海等[35]在研究中发现拟南芥在低温处理后CYP79B2的表达量呈下降趋势,本研究中,茎尖在低温处理后表达降低,与前人在拟南芥中的研究结果相似。另外,茎尖在营养生长期BrcCYP79B2-2的表达量较高,在花芽即将分化和分化时降低,这可能表示植物体内存在一种机制可以降解浓度过高的生长素,来完成花芽分化的过程。
4 结论
从普通白菜中成功克隆到CYP79B2同源基因BrcCYP79B2-2,其编码序列全长为1 554 bp,编码517 个氨基酸,在白菜不同变种中高度保守。BrcCYP79B2-2具有组织特异性,且参与对低温的响应。BrcCYP79B2-2在茎尖不同发育阶段的动态变化,说明其可能具有促进花芽分化的功能。