梨小食心虫幼虫中肠中高表达消化酶和解毒酶基因的筛选
2022-12-28吕栋标张战利冷春蒙袁向群李怡萍
吕栋标, 张战利, 冷春蒙, 袁向群, 李怡萍,*
(1. 西北农林科技大学植物保护学院, 植保资源与病虫害治理教育部重点实验室, 陕西杨凌 712100; 2. 陕西省植物保护工作总站, 西安 710003; 3. 西北农林科技大学植物保护学院, 农业部西北黄土高原作物有害生物综合治理重点实验室, 陕西杨凌 712100)
梨小食心虫Grapholitamolesta是一种重要的世界性果树害虫,广泛分布于亚洲、欧洲、美洲和大洋洲的温带与亚热带地区(Chenetal., 2020)。在我国,除西藏未见报道外,广布全国各地,尤其在东北、华北、西北、华东地区的桃、梨主产区普遍发生,危害严重,影响了果农的经济收入(冷春蒙等, 2018)。梨小食心虫在我国每年发生3~5代,以老熟幼虫在枝干裂缝、根茎基部和落叶杂草中越冬(李帅等, 2018)。越冬代成虫将卵产在树枝顶端嫩梢附近,卵孵化后蛀入嫩梢进行取食危害,造成枝条萎蔫。幼虫主要蛀食果实,排泄物会被堆积在虫孔周围,然后逐渐变黑,最后形成一大块黑斑,俗称“黑膏药”,这也是梨小食心虫取食危害的明显特征(Duetal., 2015)。梨小食心虫以幼虫钻蛀危害,化学农药不易触,其次,成虫个体小、习性隐蔽和世代重叠严重等特点,致使化学防治困难,防治效果欠佳(冷春蒙等, 2018),且化学防治的弊端“3R”问题日益突出。果实套袋是传统防治病害的有效方法之一,但均衡人力消耗和高效性的考虑,该方法迟早要淘汰。因此,需要探索绿色、高效、靶标性强的其他防治技术来解决这些问题。
昆虫中肠是昆虫消化与吸收营养物质、进行解毒代谢的主要场所,在消化酶的分泌、食物的消化和防御外源有毒物质等方面有着至关重要的作用(唐庆峰等, 2005; 叶晓冬, 2014)。胰蛋白酶(trypsin, TRY)属于丝氨酸蛋白酶,氨肽酶(aminopeptidase, APN)是水解肽链N端单个氨基酸酶,与氨肽酶同属外肽酶(exopeptidase)的还有羧肽酶(carboxypeptidase, CP),三者在昆虫肠道消化食物中起着重要的作用(赵爱平等, 2016, 2017)。昆虫在取食过程中会摄入大量的毒素,为保证昆虫正常生长发育,昆虫肠道需要解毒代谢毒素,因此,解毒酶在中肠中起着重要作用。 细胞色素P450(cytochrome P450, CYP450)、谷胱甘肽S-转移酶(glutathione S-transferase, GST)和羧酸酯酶(carboxylesterase, CarE)是3种主要的解毒酶。转录组测序可以快速获得物种的基因数据信息,包括基因挖掘、分子标记开发、基因表达分析等方面内容,为后续研究提供强大的理论支持和丰富的遗传信息(张棋麟和袁明龙, 2013)。鳞翅目昆虫如家蚕Bombyxmori、小菜蛾Plutellaxylostella、亚洲玉米螟Ostriniafurnacalis和草地贪夜蛾Spodopterafrugiperda等昆虫已通过转录组测序完成了丝氨酸蛋白酶基因的筛选和鉴定(Linetal., 2015; Guanetal., 2017; Liuetal., 2017; Hafeezetal., 2021)。近年来,也有对于梨小食心虫转录组测序的研究,如Jung和Kim(2014)对梨小食心虫性信息素腺体转录组测序,Li等(2015)利用高通量测序平台Illumina Mesiq对梨小食心虫触角转录组测序,Guo等(2017)对梨小食心虫卵、幼虫、蛹、成虫等不同虫态转录组测序,冷春蒙等(2018)利用高通量测序平台Illumina HiSeq X Ten对梨小食心虫的幼虫中肠转录组进行了测序。拥有如此多转录组数据,可以对数据深挖掘,对梨小食心虫幼虫中肠基因功能进行注释分析,从中筛选与消化和解毒代谢相关的基因。
基于我们前期对梨小食心虫幼虫中肠转录组测序及SSR分子标记的研究基础和拥有完整转录组数据,深入研究梨小食心虫消化酶和解毒酶的功能,阐释梨小食心虫取食寄主的多样性机制,本研究对梨小食心虫幼虫中肠转录组高表达的基因进行GO功能分类和KEGG代谢通路分析,筛选出具有完整开放阅读框的消化酶和解毒酶基因,将其编码的蛋白和鳞翅目其他昆虫的同源蛋白进行进化分析,并检测基因在幼虫不同龄期中肠中的表达量。研究将获得梨小食心虫消化酶和解毒酶的重要数据信息,同时为以中肠消化酶和解毒酶为靶标的新型农药研发和转基因作物提供重要的理论参考依据。
1 材料与方法
1.1 供试昆虫
本研究试虫梨小食心虫幼虫由西北农林科技大学植物保护学院害虫防治研究室提供,饲养条件和饲料配方参照杜娟(2009)的方法。 在温度26±0.5℃,相对湿度70%±10%,光周期15L∶9D (光照:6:00-21:00时,4 800 lx)的Boxun-BIC400人工气候箱,以人工饲料饲养种群。
1.2 试验试剂
Trizol提取液(赛默飞)、三氯甲烷(阿拉丁)、异丙醇(阿拉丁)、75%乙醇(阿拉丁)、反转录合成试剂盒(艾科瑞)、ChamQ SYBR qPCR Master Mix(诺唯赞)和2×Accurate Taq Master Mix(艾科瑞)等。
1.3 梨小食心虫幼虫中肠总RNA的提取与cDNA合成
将人工饲养的梨小食心虫2-5龄幼虫在体视镜下冰浴解剖中肠,并以0.15 mol/L氯化钠溶液进行多次冲洗,将各样本中肠组织液氮冷冻,保存于-80℃备用。每样本设3个生物学重复,每重复50头。采用Trizol法提取不同龄期梨小食心虫幼虫中肠总RNA,利用1.0%琼脂糖凝胶电泳和赛默飞2000分光光度计检测RNA纯度(OD260/OD280)和浓度,检测合格的RNA采用艾科瑞荧光定量反转录试剂盒合成cDNA。
1.4 梨小食心虫幼虫中肠中高表达基因分析
对本实验室前期获得的梨小食心虫4龄幼虫中肠转录组数据,采用RSEM法计算转录本的表达量,以FPKM(fragments per kilobase of exon model per million mapped fragments)值表示基因的表达水平。选取质控得到的样品转录组数据FPKM值大于15的高通量基因组序列称为高表达基因,进行GO分类和KEGG通路富集分析(Lietal., 2013)。
1.5 消化酶和解毒酶基因的筛选、鉴定与序列分析
基于1.4节功能注释信息,BLAST同源搜索从中肠转录组中筛选和鉴定高表达消化酶和解毒酶基因。从NCBI数据库中下载棉铃虫Helicoverpaarmigera、小菜蛾和草地贪夜蛾等其他鳞翅目昆虫同源基因编码的氨基酸序列,通过MEGA采用Clustal X进行多序列比对,利用邻接法(neighbour-joining method, NJ)构建系统发育树,进行1 000次bootstrap验证结果。
1.6 消化酶和解毒酶基因表达量检测
利用qRT-PCR检测1.5节筛选到的消化酶胰蛋白酶(TRY)、氨肽酶(APN)和羧肽酶(CP)以及解毒酶谷胱甘肽S-转移酶(GST)、细胞色素P450(CYP450)以及羧酸酯酶(CarE)基因在梨小食心虫2-5龄幼虫中肠中的表达量。参照张国辉和仵均祥(2015)的引物,选取梨小食心虫β-actin(GenBank登录号: JN857938)和EF-1α(GenBank登录号: KT363835)为内参基因,根据上述筛选的6种高表达基因,每种分别随机选择2个基因,利用Primer Premier 5.0软件设计qRT-PCR的引物,引物信息如表1所示。

表1 基因和引物信息Table 1 Information of genes and primers
以1.3节合成的梨小食心虫2-5龄幼虫中肠cDNA为模板,用染色法进行qRT-PCR扩增反应,LightCycler480进行检测。利用ChamQ SYBR qPCR Master Mix,反应体系(25 μL): ChamQ SYBR qPCR Master Mix (2×) 12.5 μL, 正反向引物(10 μmol/L)各1.0 μL, cDNA 2.0 μL, RNase-free ddH2O 8.5 μL。PCR扩增程序: 95℃ 30 s; 95℃ 10 s, 60℃ 60 s, 40个循环。相对定量分析采用2-ΔΔCt法。
1.7 数据分析
消化酶和解毒酶基因在梨小食心虫不同龄期的表达量数据初步整理后,利用SPSS软件(SPSS17.0,SPSS Inc.,美国)对其进行统计分析,采用单因素方差分析(one-way ANOVA)Turkey氏法进行差异显著性比较(P<0.05)。
2 结果
2.1 梨小食心虫4龄幼虫中肠转录组中高表达基因的GO功能注释和KEGG通路
GO功能分类统计(图1)表明,103 677个高表达基因主要被注释在生物学进程(biological process)、细胞组分(cellular component)和分子功能(molecular function)3个大类共41个分支。其中生物学进程中参与细胞进程(cellular process)最多(10 501个),其次是代谢进程(metabolic process)(9 548个)和单一生物体进程(single-organism process)(8 341个)。分子功能中参与结合(binding)和催化活性(catalytic activity)较多,分别为10 700和7 680个。细胞组分中参与细胞(cell)和细胞部分(cell part)最多,均为5 904个,其次是细胞器(organelle)、大分子复合物(macromolecular complex)、膜(membrane)和膜部分(membrane part),分别为4 028, 3 703, 3 586和3 357个。

图1 梨小食心虫4龄幼虫中肠转录组中高表达基因的GO功能注释Fig. 1 GO function annotation of highly expressed genes in the transcriptome of the 4th larval midgut of Grapholita molesta
KEGG通路富集分析如图2所示,共有10 846个高表达基因参与了32组子代谢通路,5类生化代谢通路。细胞进程注释最多的代谢通路是运输和分解代谢(transport and catabolism),为739个;环境信息处理(environmental information processing)注释最多的代谢通路是信号转导(signal transduction),为1 167个;遗传信息处理(genetic information processing)注释最多的代谢通路是翻译(translation),为747个;新陈代谢(metabolism)注释最多的代谢通路是碳水化合物代谢(carbohydrate metabolism),为597个;有机体系统(organismal system)注释最多的代谢通路是内分泌系统(endocrine system),为681个。

图2 梨小食心虫4龄幼虫中肠转录组中高表达基因的KEGG代谢通路Fig. 2 KEGG metabolic pathways of highly expressed genes in the transcriptome of the 4th instar larval midgut of Grapholita molestaA: 细胞进程Cellular process; B: 环境信息处理Environmental information processing; C: 遗传信息处理Genetic information processing; D: 新陈代谢Metabolism; E: 有机体系统Organismal system.
2.2 梨小食心虫4龄幼虫中肠转录组中高表达消化酶和解毒酶基因
筛选到具有完整开放阅读框的5个胰蛋白酶(TRY)基因、3个氨肽酶(APN)基因和9个羧肽酶(CP)基因都属于消化酶基因。 GmolTRY CL10148.19112, GmolTRY CL10148.19006, GmolTRY CL10148.20460, GmolTRY CL10148.17450和GmolTRY CL10148.18912具有保守的活性三联体His, Asp和Ser,同样具有RIVGG和CQGDSGGP保守的末端序列,是典型的胰蛋白酶。基于GmolTRYs和其他类群昆虫的TRYs氨基酸序列构建系统发育树(图3),发现梨小食心虫的5个TRYs分支聚类较为分散,但都至少与1个鳞翅目昆虫的TRY聚类在一支,且序列一致性很高。GmolTRY CL10148.19112和GmolTRY CL10148.17450分别与大豆食心虫LeguminivoraglycinivorellaLglyTRY(GenBank登录号: AFP74111.1)和烟草天蛾ManducasextaMsexTRY(GenBank登录号: KAG6444168.1)聚到一支。GmolTRY CL10148.19006和GmolTRY CL10148.18912与阿波罗绢蝶ParnassiusapolloPapoTRYs(GenBank登录号: CAG4992335.1和CAG5037487.1)聚到一支。

图3 邻接法构建的基于氨基酸序列的梨小食心虫4龄幼虫中肠转录组中5个GmolTRYs与其他鳞翅目昆虫胰蛋白酶(TRYs)的系统发育树(1 000次重复)Fig. 3 Phylogenetic tree of five GmolTRYs in the transcriptome of the 4th instar larval midgut of Grapholita molestaand trypsins (TRYs) from other lepidopteran insects based on amino acid sequences using neighbor-joining method (1 000 replicates)蛋白来源物种Origin species of proteins: Gmol: 梨小食心虫Grapholita molesta; Dchr: 金斑蝶Danaus chrysippus; Harm: 棉铃虫Helicoverpa armigera; Lsin: 条纹小粉蝶Leptidea sinapis; Mcon: 蓓带夜蛾Mamestra configurata; Pxut: 柑橘凤蝶Papilio xuthus; Papo: 阿波罗绢蝶Parnassius apollo; Sfru: 草地贪夜蛾Spodoptera frugiperda; Tni: 粉纹夜蛾Trichoplusia ni; Bino: 伊诺小豹蛱蝶Brenthis ino; Csup: 二化螟Chilo suppressalis; Dple: 黑脉金斑蝶Danaus plexippus; Dsac: 小蔗杆草螟Diatraea saccharalis; Pmac: 金凤蝶Papilio machaon; Obru: 冬尺蛾Operophtera brumata; Pxyl: 小菜蛾Plutella xylostella; Sexi: 甜菜夜蛾Spodoptera exigua; Aarg: 木棉虫Alabama argillacea; Onub: 欧洲玉米螟Ostrinia nubilalis; Snon: 蛀茎夜蛾Sesamia nonagrioides; Cfum: 云杉色卷蛾Choristoneurafumiferana; Hpun: 斑实夜蛾Helicoverpa punctigera; Hvir: 烟芽夜蛾Heliothis virescens; Hvir: 苜蓿夜蛾Heliothis viriplaca; Lgly: 大豆食心虫Leguminivora glycinivorella; Msex: 烟草天蛾Manduca sexta; Msep: 黏虫Mythimna separata; Tbis: 衣蛾Tineola bisselliella. 下图同The same for the following figures.
APNs(图4)系统发育树显示,梨小食心虫的3个APNs分支聚类较为分散,且都与1个鳞翅目昆虫的APN聚类在一支,且序列一致性很高。 GmolAPN CL10148.19035, GmolAPN CL10148.20703和GmolTRY CL10148.19013分别与苹淡褐卷蛾EpiphyaspostvittanaEposAPN(GenBank登录号: AAF99701.1)、二化螟ChilosuppressalisCsupAPN(GenBank登录号: RVE53189.1)和冬尺蛾OperophterabrumataObruAPN(GenBank登录号: KOB65200.1)聚到一支。

图4 邻接法构建的基于氨基酸序列的梨小食心虫4龄幼虫中肠转录组中3个GmolAPNs与其他鳞翅目昆虫氨肽酶(APNs)的系统发育树(1 000次重复)Fig. 4 Phylogenetic tree of three GmolAPNs in the transcriptome of the 4th instar larval midgut of Grapholita molestaand aminopeptidases (APNs) from other lepidopteran insects based on amino acid sequences using neighbor-joining method (1 000 replicates)蛋白来源物种Origin species of proteins: Bmor: 家蚕Bombyx mori; Ldis: 舞毒蛾Lymantria dispar; Ofur: 亚洲玉米螟Ostrinia furnacalis; Hcun: 美国白蛾Hyphantria cunea; Epos: 苹淡褐卷蛾Epiphyas postvittana; Lxyl: 木毒蛾Lymantria xylina; Pint: 印度谷螟Plodia interpunctella; Sinf: 大螟Sesamia inferens; Cinc: 黄豆银纹夜蛾Chrysodeixis includes; Cmed: 稻纵卷叶螟Cnaphalocrocis medinalis; Mbra: 甘蓝夜蛾Mamestra brassicae. 下图同The same for the following figures.
CPs系统发育树(图5)显示,梨小食心虫的9个GmolCPs分支聚类较为分散,但都至少与1个鳞翅目昆虫的CP聚类在一支,且序列一致性很高。 GmolCP CL10148.18096, GmolCP CL10148.19435, GmolCP CL10148.16225和GmolCP CL10148.11888分别与二化螟CsupCPs(GenBank登录号: RVE46147.1, RVE46146.1, RVE53886.1和RVE53889.1)聚到一支。 GmolCP CL10148.19435, GmolCP CL10148/19598, GmolCP CL10148.23889和GmolCP CL10148.19821分别与阿波罗绢蝶PapoCPs(GenBank登录号: CAG4972060.1, CAG4967861.1, CAG5016938.1和CAG5004946.1)聚到一支。 GmolCP CL10148.15735与另外8个GmolCPs没有聚到同一分支,和其他鳞翅目昆虫的CPs单独聚到一支。
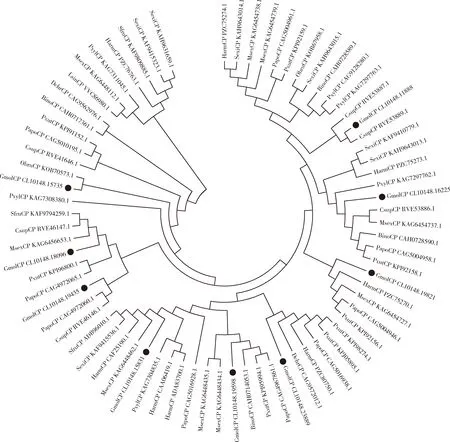
图5 邻接法构建的基于氨基酸序列的梨小食心虫4龄幼虫中肠转录组中9个GmolCPs与其他鳞翅目昆虫羧肽酶(CPs)的系统发育树(1 000次重复)Fig. 5 Phylogenetic tree of nine GmolCPs in the transcriptome of the 4th instar larval midgut of Grapholita molestaand carboxypeptidases (CPs) from other lepidopteran insect based on amino acid sequences using neighbor-joining method (1 000 replicates)
筛选到具有完整开放阅读框的11个谷胱甘肽S-转移酶(GST)基因、13个细胞色素P450(CYP450)基因和8个羧酸脂酶(CarE)基因都属于解毒酶基因。GSTs系统发育树(图6)显示,梨小食心虫11个GSTs聚类较为集中,GmolGST CL10148.20001, GmolGST CL10148.20002和GmolGST CL10148.20240聚到一支,同时与苹果蠹蛾CydiapomonellaCpomGST(GenBank登录号: ARM39000.1)聚为一支。 GmolGST CL10148.18663, GmolGST CL10148.19812, GmolGST CL10148.20710, GmolGST CL10148.19617, GmolGST CL10148.19740和GmolGST CL10148.20112分别与苹果蠹蛾CpomGSTs(GenBank登录号: ARM39003.1, ARM39002.1, ARM39006.1, AYX80656.1, ARM39009.1和AYX80655.1)聚到一支。 GmolGST CL10148.18801和GmolGST CL10148.16901分别与梨小食心虫GmolGSTs(GenBank登录号: AVX32544.1和AVZ44716.1)聚到一支。

图6 邻接法构建的基于氨基酸序列的梨小食心虫4龄幼虫中肠转录组中11个GmolGSTs与其他鳞翅目昆虫谷胱甘肽S-转移酶(GSTs)的系统发育树(1 000次重复)Fig. 6 Phylogenetic tree of 11 GmolGSTs in the transcriptome of the 4th instar larval midgut of Grapholita molestaand glutathione S-transferases (GSTs) from other lepidopteran insect species based on amino acid sequences using neighbor-joining method (1 000 replicates)蛋白来源物种Origin species of proteins: Cpom: 苹果蠹蛾Cydia pomonella; Gpyl: 桑绢野螟Glyphodes pyloalis; Hvit: 黄野螟Heortia vitessoides; Slit: 海灰翅夜蛾Spodoptera littoralis; Hass: 烟青虫Helicoverpa assulta; Aper: 柞蚕Antheraea pernyi; Aips: 小地老虎Agrotis ipsilon; Cfum: 云杉卷叶蛾Choristoneura fumiferana; Mtro: 杨小舟蛾Micromelalopha troglodyta; Gmel: 大蜡螟Galleria mellonella. 下图同The same for the following figures.
CYP450s系统发育树(图7)显示,梨小食心虫13个CYP450s聚类较为集中,GmolCYP450 CL10148.27307和GmolCYP450 CL10148.23751聚为一支。 GmolCYP450 CL10148.180049, GmolCYP450 CL10148.21072, GmolCYP450 CL10148.20009, GmolCYP450 CL10148.19051, GmolCYP450 CL10148.19496和GmolCYP450 CL10148.19528分别与苹果蠹蛾CpomCYP450s(GenBank登录号: AZJ25110.1, AXP17117.1, AXP17131.1, AZJ25111.1, AXP17178.1, AXP17211.1, AZJ25098.1和AXP17158.1)聚为一支。GmolCYP450 CL10148.25748和GmolCYP450 CL10148.21217分别与小菜蛾PxylCYP450(GenBank登录号: KAG7296067.1)和茶小卷叶蛾AdoxophyeshonmaiAhonCYP450(GenBank登录号: BBE49547.1)聚为一支。

图7 邻接法构建的基于氨基酸序列的梨小食心虫4龄幼虫中肠转录组中13个GmolCYP450s与其他鳞翅目昆虫细胞色素P450(CYP450s)的系统发育树(1 000次重复)Fig. 7 Phylogenetic tree of 13 GmolCYP450s in the transcriptome of the 4th instar larval midgut of Grapholita molestaand cytochrome P450s (CYP450s) from other lepidopteran insects based on amino acid sequences using neighbor-joining method (1 000 replicates)蛋白来源物种Origin species of proteins: Zfil: 六星灯蛾Zygaena filipendulae; Mcin: 庆网蛱蝶Melitaea cinxia; Prap: 菜粉蝶Pieris rapae; Ahon: 茶小卷叶蛾Adoxophyes honmai; Bman: 野桑蚕Bombyx mandarina; Eelu: 烟草粉螟Ephestia elutella. 图8同The same for Fig. 8.
CarEs系统发育树(图8)显示,梨小食心虫的8个CarEs分支聚类较为分散,但都至少与1个鳞翅目昆虫的CarE聚类在一支,且序列一致性很高。 GmolCarE CL10148.19098, GmolCarE CL10148.15194和GmolCarE CL10148.9606分别与苹果蠹蛾CpomCarEs(GenBank登录号: AKP92859.1, AMB19667.1和AKP92857.1)聚到一支。 GmolCarE CL10148.15334, GmolCarE CL10148.15735和GmolCarE CL10148.20762分别与伊诺小豹蛱蝶BrenthisinoBinoCarE(GenBank登录号: CAH0729058.1), 冬尺蛾ObruCarE(GenBank登录号: KOB70573.1)和二化螟CsupCarE(GenBank登录号: AIY69075.1)聚为一支。GmolCarE CL10148.11834和其他的GmolCarEs没有聚到一支,和其他鳞翅目昆虫的CarEs聚成独立的小分支。
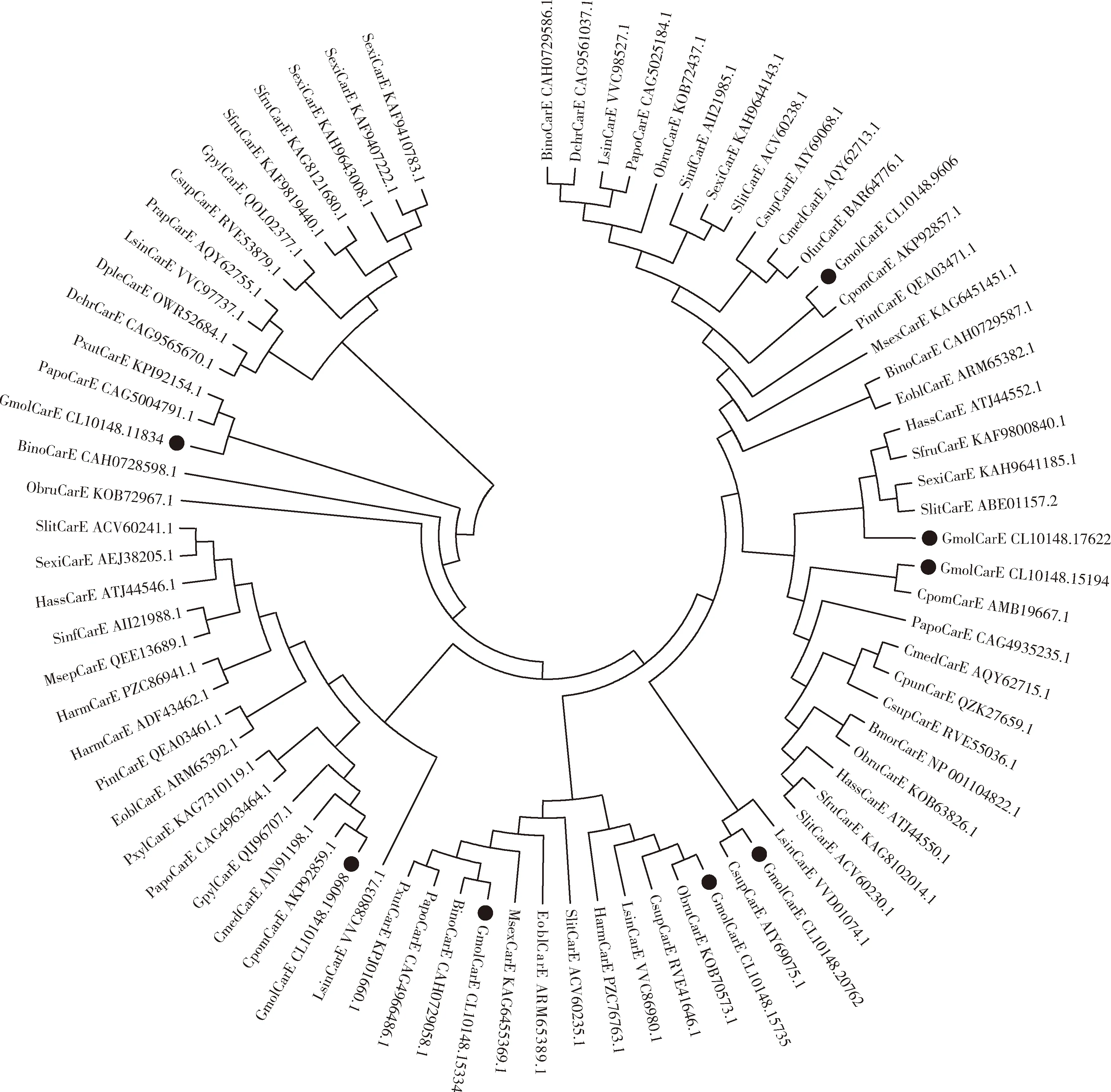
图8 邻接法构建的基于氨基酸序列的梨小食心虫4龄幼虫中肠转录组中8个GmolCarEs与其他麟翅目昆虫羧酸酯酶(CarEs)的系统发育树(1 000次重复)Fig. 8 Phylogenetic tree of eight GmolCarEs in the transcriptome of the 4th instar larval midgut of Grapholita molestaand carboxylesterases (CarEs) from other lepidopteran insects based on amino acid sequences using neighbor-joining method (1 000 replicates)蛋白来源物种Origin species of proteins: Eobl: 茶尺蠖Ectropis obliqua; Cpun: 桃蛀螟Conogethes punctiferalis.
2.3 梨小食心虫4龄幼虫中肠转录组中高表达消化酶和解毒酶基因的表达量
消化酶和解毒酶基因在梨小食心虫不同龄期幼虫中肠的表达量差异显著,基因表达量随龄期呈现先增长后下降的变化趋势,表达量均在4龄幼虫期最高,消化酶基因各幼虫龄期表达量依次为4龄>3龄>2龄>5龄,解毒酶基因各幼虫龄期表达量依次为4龄>3龄>5龄>2龄(图9)。消化酶基因在梨小食心虫各幼虫龄期的表达量特点不同,4龄时,TRY基因的表达量为2龄时的4倍左右,APN基因和CP基因的表达量均为2龄时的3倍左右(P<0.05);3龄时,TRY基因、APN基因和CP基因的表达量均为4龄时的一半左右(P<0.05)(图9: A-C)。解毒酶基因在梨小食心虫各幼虫龄期的表达量特点不同,4龄时,CYP450基因、GST基因和CarE基因的表达量均为2龄时的2.5倍左右;CYP450基因(CL10148.19323)在各幼虫龄期表达量差异显著(P<0.05),CYP450基因(CL10148.19496)在3龄和4龄幼虫中的表达量无显著差异(P>0.05)。 GST基因和CarE基因在3龄和4龄幼虫中的表达量均差异显著(P<0.05)(图9: E, F)。

图9 消化酶(A-C)和解毒酶(D-F)基因在梨小食心虫不同龄期幼虫中肠中的表达量Fig. 9 Expression levels of digestive enzyme (A-C) and detoxification enzyme (D-F) genes in the midgut of Grapholita molesta larvae at different instarsA: 胰蛋白酶Trypsin (TRY); B: 氨肽酶Aminopeptidase (APN); C: 羧肽酶Carboxypeptidase (CP); D: 细胞色素P450 Cytochrome P450 (CYP450); E: 谷胱甘肽S-转移酶Glutathione S-transferase (GST); F: 羧酸脂酶Carboxylesterase (CarE). 图中数据分别为3组重复实验数据的平均值±标准误; 柱上不同小写字母表示基因在不同龄期间的相对表达量差异显著(P<0.05, ANOVA, Tukey氏HSD)。Data in the figure are means±SE. Different lowercase letters above bars indicate significant difference in the relative expression levels of genes between different instars (P<0.05, ANOVA, Tukey’s HSD).
3 讨论
本研究对前期梨小食心虫幼虫中肠转录组高表达基因的分析发现,GO功能分类中参与细胞进程、代谢进程和催化活性的基因较多(图1)。KEGG代谢通路在细胞进程中注释最多是运输和分解代谢,新陈代谢中注释最多是碳水化合物代谢通路(图2)。大量高表达基因参与细胞进程与代谢和运输与分解代谢,说明梨小食心虫幼虫在发育过程中需要大量的蛋白质和能量,因此蛋白质和能量代谢较为旺盛。物质代谢的主导地位说明此时的梨小食心虫幼虫有着较强的消化吸收能力,主要用于梨小食心虫的消化和生长,与前期的研究结果(冷春蒙等, 2018)相同。但仍有一些注释到非主要功能的基因,说明为了维持肠道的稳定,也有很多细胞生物功能的基因参与。
通过高表达功能基因筛选,本研究得到完整开放阅读框的消化酶基因17个(5个TRY基因、3个APN基因和9个CP基因)以及解毒酶基因32个(11个GST基因、13个CYP450基因和8个CarE基因)。与其他鳞翅目昆虫相比获得的数量相对较少,可能是因为所取部位仅为幼虫中肠,而头、外表皮、脂肪体、马氏管和触角等部位也存在与此类相关的高表达基因。通过将消化酶和解毒酶分别与鳞翅目其他昆虫同源蛋白氨基酸序列构建的系统发育树,发现梨小食心虫消化酶和大豆食心虫、苹淡褐卷蛾、烟草天蛾和阿波罗绢蝶的同源蛋白亲缘关系较近(图3-5),解毒酶和苹果蠹蛾、茶小卷叶蛾、小菜蛾、二化螟的同源蛋白亲缘关系较近(图6-8)。由于转录组数据需要和NCBI基因数据库(https:∥www.ncbi.nih.gov)中参考序列比对,虽然烟草天蛾、阿波罗绢蝶、小菜蛾和二化螟虽然都属鳞翅目,但分别属于天蛾科、绢蝶科、菜蛾科和螟蛾科,可能由于国内外对烟草天蛾、阿波罗绢蝶、小菜蛾和二化螟的研究较多,NCBI数据库数据较为丰富,所以梨小食心虫和它们亲缘关系较近。消化酶与同源分支聚类较为分散,可能因为昆虫的消化过程较为复杂,消化酶基因为一个庞大的家族,本研究获得的基因可能属于不同的家族成员。GSTs和CYP450s与同源分支聚类较为聚中,可能因为GSTs和CYP450s本就属于重要的解毒酶,本研究筛选的是高表达基因,所以GSTs和CYP450s聚类较为集中。
基于对梨小食心虫GO功能分类和KEGG代谢通路的分析,本研究筛选出高表达的解毒酶和消化酶基因,并对各类代表性基因在不同龄期进行表达量检测,消化酶基因表达量变化趋势和赵爱平等(2016)对胰蛋白酶基因的研究结果相同,表达量差异显著,依次为4龄>3龄>2龄>5龄(图9)。赵爱平等(2016)对梨小食心虫幼虫中肠蛋白酶活性进行了测定,发现酶活性变化趋势和胰蛋白酶基因表达趋势一致。美国白蛾氨肽酶基因在各龄期表达量变化趋势(Zhangetal., 2017)和本研究结果一致。家蚕羧肽酶部分基因在5龄幼虫中肠中高表达,大螟和二化螟的氨肽酶基因在3龄幼虫中肠中高表达(王兴云等, 2012; 朱玉苹等, 2016; Wangetal., 2017; 展恩玲, 2018)。本研究发现梨小食心虫部分氨肽酶基因在2龄幼虫中肠中高表达,可能是昆虫物种不同而导致的差异。本研究中解毒酶基因在各幼虫龄期的表达量变化趋势依次为4龄>3龄>5龄>2龄(图9)。白琪等(2018)研究发现二化螟两种CYP450基因在幼虫中随龄期变化的表达量同样是先增长后下降的趋势;任娜娜等(2015)发现在小菜蛾4龄幼虫中羧酸酯酶基因的表达量最高,显著高于其他龄期的;刘月庆(2017)发现粘虫各龄期幼虫的CYP450基因表达量差异显著,5龄和6龄时达到最大值。本研究结果与上述研究中解毒酶基因的表达量变化趋势一致。
本研究通过GO和KEGG数据库成功筛选和验证部分高表达消化酶和解毒酶基因,研究结果为鳞翅目其他近缘物种昆虫的肠道转录组的研究提供了重要的数据信息,对鳞翅目卷蛾科非模式昆虫的研究有重要的推动意义,同时为研究以肠道为靶标的新型农药和转基因作物提供了重要的理论参考依据。但要明确消化酶和解毒酶基因的具体功能,还需后续深入研究。
