Quantification and Characterization of Rubber from Primary Laticifers of Hevea brasiliensis
2022-12-16YAOXingchengTUHanqiZHOUJunCHENXianhongCHENQingLINWeifuWANGJun
YAO Xingcheng, TU Hanqi, ZHOU Jun, CHEN Xianhong, CHEN Qing, LIN Weifu, WANG Jun*
Quantification and Characterization of Rubber from Primary Laticifers of
YAO Xingcheng1,2, TU Hanqi1,2, ZHOU Jun1,2, CHEN Xianhong1,2, CHEN Qing1,2, LIN Weifu1,2, WANG Jun1,2*
1. Rubber Research Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan 571101, China; 2. Key Laboratory of Biology and Genetic Resources of Rubber Tree, Ministry of Agriculture and Rural Affairs, Haikou, Hainan 571101, China
contains two types of laticifers, primary and secondary. The former distributes in the leaf and bark of stems with primary growth, and the latter distributes in the bark of stems with secondary growth. The commercial rubber is extracted from the latex of secondary laticifers and much is known to this rubber. However, little is known to the rubber in primary laticifers as it has not been exploited. This study focused on the rubber from primary laticifers from young shoots of grafted plants. We evaluated the rubber yield-related factors, including shoot biomass, biomass partitioning, and rubber content, to determine rubber yield potential. In addition, we characterized the physical and chemical properties and vulcanization behavior of raw rubbers. Results showed that clones could affect shoot biomass, biomass partitioning, and rubber content. On average, biomass partitioning value for the leaf, bark and wood was 55.5%, 10.5%, and 34.0%, respectively. Rubber content in the leaf and bark was 13.7 and 35.6 mg/g dry weight, respectively. The rubber particle average size of the latex from primary laticifers was 0.36 μm, smaller than that from secondary laticifers. Rubber from primary laticifers had a smaller molecular weight than that from conventionally tapped mature trees. The nitrogen content of rubber from primary laticifers was 0.56%, higher than that of conventional rubber (0.50%). The plasticity retention index (PRI) of rubber from primary laticifers was 89%, higher than that of conventional rubber (80%). However, the Mooney viscosity and initial plasticity (0) of the rubber from primary laticifers was 66 and 26, respectively, lower than that of conventional rubber (101 and 50, respectively). The volatile matter content and the ash content of rubber from primary laticifers was 0.50% and 0.38%, respectively, the same as that of conventional rubber. The scorching10and optimal vulcanization90times of the rubber from primary laticifers was 2.00 and 19.12 min, respectively, which was lower than conventional rubber (2.47 and 25.08 min, respectively). Comparing to conventional rubber, rubber from primary laticifers exhibited a lower minimum torque (L) value (0.22 dN·m), but a higher maximum torque (H) value (8.25 dN·m). By analyzing three clones, the results suggest that the rubber yield from primary laticifers from young shoots could be affected by shoot biomass, biomass partitioning and rubber contents. This rubber yield related results would provide a foundation for future work on breeding, germplasm development and cultivation practices. The results on rubber property showed that the rubber from primary laticifers was comparable to conventional rubber except for Mooney viscosity and0, suggesting that the rubber from primary laticifers could not be used as standard rubber but general rubber material.
; natural rubber; primary laticifers; physical and chemical properties
1 Introduction
(Willd. ex A. Juss.) Müll. Arg., also known as the Para rubber tree, is a tropical tree native to the tropical regions of South America and belongs to the family. This species is found in Amazonia and adjoining areas and is the sole source of commercial natural rubber (NR), which is used to create more than 52 000 products directly or indirectly[1]. Because of its economic importance, more than 10 million hectares ofhave been planted worldwide, with production reaching 13.5 million tons in 2017, of which 91% was produced by Asian countries[2-3]. China consumes a large amount of NR but produces very little, as the strict climate requirements oflimit its planting area and rubber yield. Furthermore, certain areas of China belong to non-traditional rubber-growing areas, with extreme environmental conditions such as low temperatures and typhoons[4-5].
latex is biosynthesized and stored in laticifers, which are specialized cells within the tree.contains two types of laticifers: primary and secondary. Primary laticifers appear along with primary growth and are mainly distributed in leaves and young stems. For the young plants within two years, the relationship between laticifer differentiation and shoot growth can be identified in the new growing shoots, where only primary laticifers are present in the youngest four whorls (also called flush or extension units), while secondary laticifers will not emerge until the fifth whorl[6-7]. For the plants more than three years, the green shoots or stems with leaves contain primary laticifers, and the brown stems without leaves contain secondary laticifers.
Therubber from secondary laticifers is commercially exploited by tapping trunks of mature trees. The economic importance of natural rubber has promoted many studies on the rubber from secondary laticifers, including latex biogenesis, rubber yield, chemical and physical properties. Primary laticifers are not tapped in production practice, therefore, less effort has been devoted to the rubber from primary laticifers. A few studies have been focused on comparisons of primary and secondary laticifers, including laticifer morphology and transcriptome[8], laticifer differentiation[9], lutoid proteomics[10], to better understand the latex biogenesis in secondary laticifers. The rubber from primary laticifers extracting young seedlings was also compared with that from mature plants for understanding the mechanisms controlling molecular structure and molecular weight[11-12].
Rubber production from secondary laticifers in mature rubber trees requires a long non-productive period (about 6‒9 years) and production cycle (about 30 years). Extracting the rubber in primary laticifers from shoots of young plants may provide an alternative way for rubber production. According to this production mode, shoots can be harvested by pollarding the plants and then used for rubber extraction. However, system knowledge on rubber from primary laticifers is lack.
In the present study we investigated the quantification and characterization of rubber in primary laticifers in the shoots of. We also evaluated rubber-related factors including biomass, biomass partitioning, and rubber contents. The technical properties of the rubber were also investigated.
2 Materials and methods
2.1 Plant materials
Grafted plants of various clones (CATAS 7-20-59, CATAS 7-33-97, and CATAS 879) were planted in 2015 with a spacing of 0.6 m×0.6 m in a field on a farm in Danzhou (19°50′N, 109°48′E, at an elevation of 158 m), Hainan, China. The plants were pollarded about 0.5 m above the ground in November 2016. After six months of growth, young shoots that grew from the remaining stumps were harvested to determine the biomass, biomass partitioning, and rubber content.
2.2 Methods
2.2.1 Biomass determination and biomass partitioning Fifteen plants were collected from one replication, with four replications for each clone. The plant materials were air-dried and separated into leaves and stems, while the bark tissue was pared manually from the stems. The leaf, bark, and wood were oven-dried at 60℃ to a constant weight and then weighed. The partitioning data are expressed as a percentage of the total dry weight.
Agronomic data were subjected to analysis of variance using SPSS statistics 20 to determine the cloning effects. Mean data were separated using Duncan’s new multiple range test at=0.05.
2.2.2 Rubber content and yield determination The oven-dried leaf and bark were ground in a mill fitted with a 1 mm mesh screen. Then, the rubber content in the leaf and bark was determined by a modified organic solvent based gravimetric method from Black.[13], as described by Suchat.[14].
Approximately 5.0 g of ground sample was placed in an extraction bag and placed into a Soxhlet apparatus (500 mL), which contained a condenser on the top and a round bottom flask, and the apparatus was placed in a thermostatic water bath to reflux. The sample was first extracted with 480 mL of acetone for 30 h at 65℃, and then with 480 mL of cyclohexane for 30 h at 90℃. The extracted cyclohexane solvent was filtrated through a paper filter that was re-wetted with fresh cyclohexane and placed onto a pre-weighed aluminum pan. The filtrate volume was increased to 480 mL with fresh cyclohexane and then evaporated in a fume hood. After evaporating until dry, the pans were oven-dried at 60℃ for 5 h and then cooled in a desiccator, and then the pans were weighed immediately after cooling. An empty extraction bag was placed into the Soxhlet apparatus, which underwent the same procedure, to serve as a blank assay. In the blank assay, the final pan weight value, after subtracting the initial pan weight, was used as the corrected value.The rubber content was calculated according to the following formula: [final pan weight−initial pan weight−correct value]/sample weight. Five replicates were carried out for each plant tissue, and the mean was calculated as the putative rubber content. The rubber yield was derived from the rubber content multiplied by the biomass and biomass partitioning.
2.2.3 Rubber particle size and molecular weight analysis The latex in the primary laticifers was collected by cutting the petioles and green stems from the second to fourth whorls, and the material was transferred into a tube containing an ice-cold buffer (10 mmol/L of Tris HCl, 100 mmol/L of sucrose, pH 7.5). The rubber particle size was determined using a LA 960 laser diffraction particle size analyzer (Horiba, Japan) according to the manufacturer’s directions.
The latex in the primary laticifers was collected from the cuts and was coagulated in a petri dish with 3% acetic acid. The dried rubber was then completely dissolved in tetrahydrofuran (THF) and syringe-filtered through a 1.0 μm glass microfiber GF/A filter. Molecular weights and distributions were determined by gel permeation chromatography (GPC) using an Agilent 1260 instrument with a refractive index detector. The system was equipped with two columns (waters Styragel HR2 7.8 mm× 300 mm, and waters Styragel HR5E 7.8 mm× 300 mm), with a flow rate of 1 mL/min and a column temperature of 35℃.
The conventional latex from the secondary laticifers was collected from tapped mature trees (CATAS 7-33-97, nine years old), and analyzed according to particle size and molecular weight.
2.2.4 Rubber characterization (1) Rubber sample collection and preparation. The latex from primary laticifers was collected from the cuts, and the latex from the three clones was mixed during collection. Conventional rubber was obtained by collecting the latex from mature trees with a tapping age of eight years (CATAS 7-33-97) by conventional tapping (d/4, 1/2S). The latex was then coagulated with 3% acetic acid, and the dried raw rubber was homogenized in accordance with the ISO 1795∶2007 standard.
(2) Measurement of nitrogen content, ash content, and volatile matter content. Nitrogen content (%) was determined with an automatic azotometer (ATN-1100), according to the ISO 1656:1996 standard. The ash content (%) was determined by the ISO 247:2006 standard, while the volatile-matter content (%) was determined by the ISO 248:2005 standard.
(3) Characterization of raw rubber. Mooney viscosity was determined using a shearing disc viscometer (UM-2050, U-CAN, Taiwan, China) according to the ISO 289-1:2005 standard, with 1 min of preheating and 4 min of heating at (100±0.5)℃. The initial plasticity (P0) and the plasticity after oven aging for 30 min (P30) at (140±0.5)℃ were measured using a Mark IV Wallace Rapid Plastimeter. The test sheets were 13 mm in diameter and 3.0 mm in height. PRI was calculated by:
PRI = (P30/ P0)×100%.
(4) Characterization of vulcanizates. The vulcanized rubber formula was as follows: 100 phr of sample, 3.5 phr of sulfur, 6 phr of ZnO, 0.5 phr of stearic acid, and 0.5 phr of 2-mercaptobenzothiazole. The vulcanized characteristics of the compounds were measured at 145℃ using a rotorless vulcanizer (MRD-2000, Alpha) according to the ISO 6502:1991 standard.
3 Results and discussion
3.1 Main factors that affect rubber yield from young shoots
Our results showed that the average shoot biomass was 78.6 g per plant for the three studied clones. The shoot biomass among the three clones was significantly different, and CATAS 879 had a higher value than the other two clones (Tab. 1). The shoot biomass values of CATAS 7-20-59 and CATAS 7-33-97 were similar. The leaf, bark, and wood partitioning values were significantly different among the three clones. On average, the shoot partitioned biomass was 55.5% leaf, 10.5% bark, and 34.0% wood.
Our earlier study confirmed that the cyclohexane extract obtained by acetone extraction from the tissue of youngshoots was cis-1,4-polyisoprence[15]. Therefore, solvent extraction was used to determine the rubber content intissue. Because primary laticifers are primarily distributed in leaf and bark, rubber contents in both tissues were analyzed. Our results showed that the rubber contents in the bark and leaf varied among the clones. For example, CATAS 879 had the highest rubber contents in the bark and leaf, while the lowest contents in leaf and bark were found in CATAS 7-20-59 and CATAS 7-33-97, respectively. The cloning effect on rubber content also indicated that rubber biosynthesis in the primary laticifers was genetically controlled. Furthermore, we observed that the average rubber content in the bark was 35.6 mg/g bark dry weight, which was much higher than in the leaf (13.7 mg/g leaf dry weight). According to an early study, the number of primary laticifers in the bark was much greater than that in the leaf[7]. Therefore, higher rubber content in the bark would attribute to more laticifers in the bark tissue.
The rubber yield extracted from primary laticifers in young shoots was mainly dependent on aerial biomass and its rubber content. Rubber yield from different plant parts (leaf, bark, or stem) was evaluated (Tab. 1), and we observed that the rubber yield in the shoot was different among the clones. This was attributed to the differences in biomass, biomass partitioning, and rubber content. CATAS 879 presented the highest rubber yield (1.14 g per plant, the sum of rubber yield from leaf and bark), because of its increased biomass and rubber contents in the leaf and bark. Moreover, the average rubber yield in the leaf and bark was 0.59 and 0.30 g per plant, respectively. Because the rubber produced in the stems was solely from the bark tissue, the rubber yield from the stems was the same as the bark. Although the rubber contents in the leaf (13.7 mg/g) were lower than in the bark (35.6 mg/g), the rubber yield from the leaf (0.59 g per plant) was much higher than in the bark (0.30 g per plant). Thus, the amount of leaf partitioning (55.5%), which was more than five times that of the bark (10.5%), added significantly to the rubber yield from the leaf. This indicated that an increase in leaf biomass would effectively improve rubber yield. Therefore, methods to increase leaf biomass have been suggested for cultivation.
The rubber yield produced by the shoot was quite low. Using the clone with the highest values, CATAS 879, and assuming a biannual harvest, rubber yield could reach 2.28 g/plant/year. Under a stock seedling planting density (60 cm× 30 cm), the estimated yield could reach 127 kg/hm2per year. Considering these assumptions, this theoretical yield would not be comparable to a yield obtained by conventional tapping. On Hainan Island, China, the average rubber yield for production years (from the first tapping year to the last) is about 900 kg/hm2per year, using conventional tapping. When the entire production cycle (from the planting year to the replanting year) is considered, the average rubber yield is 600 kg/hm2per year. Nevertheless, a higher yield clone could be obtained, as more clones are studied. Furthermore, cultivation techniques, such as planting density, irrigation, and fertilization, may also affect and improve rubber yield, and require further study. Genetics may also play a role in biomass, biomass partitioning, and rubber content, and through plant breeding or genetic engineering, plants with a higher rubber yield may be developed. In fact, the rubber yield of, which also produces latex in primary laticifers, was increased by different breeding methods[16]. After four selection cycles, the rubber yield ofincreased from 0.15 to 0.22 g/plant[17].
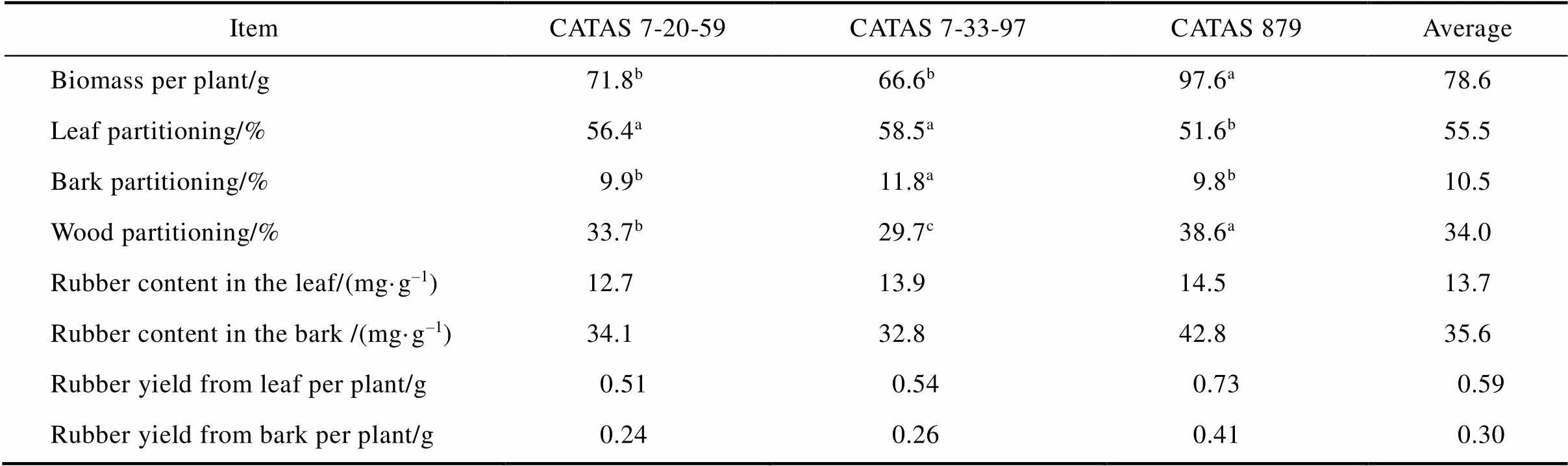
Tab. 1 Factors related to rubber yield in the different clones
Note: Values in a row followed by the same letter were not significantly difference (<0.05).
3.2 Rubber characterization
3.2.1 Rubber molecular weight and particle size analysis The rubber from primary laticifers showed a slight bimodal molecular weight distribution for the three clones (Fig. 1), and the number average molecular weight (n) and the weight average molecular weight (w) values were similar among the three clones (Tab. 2). However, the conventional rubber from tapped mature trees showed an apparent bimodal molecular weight distribution (Fig. 1). As shown in Tab. 2, the rubber from primary laticifers exhibited a smaller molecular weight compared to conventional rubber.
The rubber particle size distributions of the latex from primary laticifers among the different clones were compared. We observed that the three clones did not affect the rubber particle sizing of the latex from primary laticifers, as the particle size distributions of the three clones were almost identical (Fig. 2). The latex from primary laticifers showed an average particle size of about 0.36 μm, with a distribution ranging from 0.09 to 0.88 μm. More than 50% of the particles were in 0.3–0.45 μm in size. The rubber particles from the primary laticifers ofwere similar to[18], which only contains primary laticifers. This may indicate a similarity in the primary laticifers between the two species. The rubber particles in the latex from primary laticifers were smaller in size, when compared to the particles in conventional latex from tapped mature trees (Fig. 2). According to previous studies, rubber particle size can significantly influence molecular weight[19]and the molecular weight of large rubber particles (diameter>0.25 μm) increases with increasing particle size[20]. In addition, rubber from three-month seedlings, which only containing primary laticifers, was comprised of a few branched molecules[12]. Therefore, the lower molecular weight of rubber from primary laticifers may be caused by smaller particle sizing and/or decreased branching of the molecules, compared to conventional rubber from secondary laticifers.
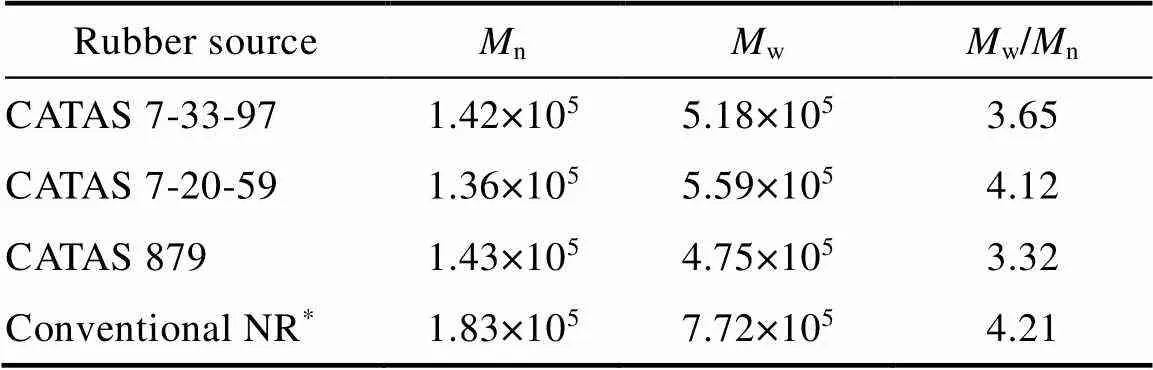
Tab. 2 Molecular characterization of rubbers from primary laticifers and conventional NR
Note: *From tapped mature trees of CATAS 7-33-97.

Fig. 1 Molecular weight distribution of the rubbers from primary laticifers and conventionally tapped trees

A: the distribution of the three clones almost overlapping; B: clone CATAS 7-33-97.
3.2.2 Characterization of raw rubber Tab. 3 shows a comparison between the rubber of primary laticifers and conventional rubbers, in terms of technological properties. The nitrogen content (N%) in the rubber from primary laticifers was 0.56%, while that of conventional rubber was 0.51%. Nitrogen in natural rubber is associated with the presence of proteins and amino acids; thus, higher N% indicated the presence of more proteins and amino acids in the rubber from primary laticifers. The ash content in the rubber from primary laticifers was 0.38%, which was the same as conventional rubber. The volatile matter content of rubber from primary laticifers and conventional rubber was identical (0.50%). The nitrogen, ash, and volatile matter content values for the rubber from primary laticifers were within the SMR specification limits.
Mooney viscosity and0are related to the chain length of polyisoprene, while0indicates the microstructure of the raw rubber[21]. When the0of rubber is less than 30 units, the rubber is considered too soft[22]. In this study, the0value of the rubber from primary laticifers was only 26, which was less than the acceptable value (30) for many standard rubber categories. Therefore, this indicated that the rubber from primary laticifers did not meet the0requirement for standard natural rubbers. The Mooney viscosity of the rubber from primary laticifer was 66, which was less than that of conventional rubber (101). This was attributed to the lower molecular weight of the rubber from primary laticifers. However, the PRI value of the rubber from primary laticifers was 89%, which was higher than that of conventional rubber (80%). PRI is used to evaluate the resistance of a material to thermo- oxidation[23]; therefore, the rubber from primary laticifers may have a higher resistance to thermo- oxidation than conventional rubber. In addition, non-rubber components such as proteins and amino acids play a role in antioxidation. Thus, the increased number of proteins and amino acids in the rubber from primary laticifers, as indicated by a higher N%, may suggest more antioxidant content, which could explain the higher PRI. Above all, the rubber from primary laticifers may not be used as standard rubber, but it could be exploited as general rubber material or supplementary material.
3.2.3 Vulcanization characteristics The characteristics of vulcanizates are presented in Tab. 4. The scorching T10and optimal vulcanization T90times of the rubber from primary laticifers were 2.00 and 19.12 min, respectively, which was lower than conventional rubber (2.47 and 25.08 min, respectively). This indicated that the rubber from primary laticifers had a higher vulcanization rate. The rubber from primary laticifers also had a lower minimum torque (L) value (0.22 dN·m), but a higher maximum torque (H) value (8.25 dN·m). The rubber from primary laticifers exhibited a higherH–Lvalue (8.03 dN·m) than conventional rubber (5.49 dN·m), indicating a higher degree of crosslinking in the vulcanized rubber from primary laticifers.

Tab. 3 Technological properties of rubber from primary laticifers and conventional NR

Tab. 4 Vulcanization characteristics of rubber from primary laticifers and conventional NR
4 Conclusions
In this study, factors related to rubber yield, variations in biomass, biomass partitioning, and rubber contents in plant tissues were observed among clones, and the results showed a genetic effect. The rubber yield related results would provide a foundation for future work on breeding, germplasm development and cultivation practices. The characteristics of rubber from primary laticifers were considered acceptable, except for the low molecular weight and0values. Therefore, the rubber from primary laticifers may not be entirely comparable with conventional rubber, and it could not be used as standard rubber; however, it could be utilized as a feedstock for the production of high value specialty polymer derivatives analogous to those produced from conventional rubber. In conclusion, this study confirmed the potential of exploiting rubber from primary laticifers fromshoots. However, the production of NR fromshoots will only be possible with significantly improved rubber yield.
[1] AGWU A E. Enhancing natural rubber () production through extension service delivery in southwest agricultural zone of Nigeria[J]. Journal of Agriculture, Food and Environment and Extention, 2006, 5(2): 7-16.
[2] IRSG. Rubber statistical bulletin[R/OL]. (2019-09-30)[2019-11-01]. http://www.rubberstudy.com/documents/WebSiteData_ Aug2019.pdf.
[3] YAO X C, CHEN X H, WANG J, ZHOU J, CAI M D, LIN W F. Effect of clonal rootstocks on the growth and yield of hevea rubber[J]. Journal of Rubber Research, 2017, 20: 203-212.
[4] HUANG Z D, PAN Y Q. Rubber cultivation under climatic stresses in China[C]//Natural Rubber: Biology, cultivation and Technology. Amsterdam: Elsevier, 1992.
[5] PRIYADARSHAN P M. Biology of hevea rubber[M]Berlin: Springer, 2017: 131.
[6] HAO B Z, WU J L. Laticifer differentiation in: induction by exogenous jasmonic acid and linolenic acid[J]. Annals of Botany, 2000, 85: 37-43.
[7] 吴继林, 谭海燕, 曾日中, 郝秉中. 巴西橡胶树初生乳管分化与苗生长的关系[J]. 热带作物学报, 2000, 21(4): 1-6.
WU J L, TAN H Y, ZENG R Z, HAO B Z. Primary laticifer differentiation ofin relation to shoot growth[J]. Chinese Journal of Tropical Crops, 2000, 21(4): 1-6. (in Chinese)
[8] TAN D, HU X, FU L, KUMPEANGKEAW A, DING Z, SUN X, ZHANG J. Comparative morphology and transcriptome analysis reveals distinct functions of the primary and secondary laticifer cells in the rubber tree[J]. Scientific Reports, 2017, 7: 3126.
[9] LOH S C, THOTTATHIL G P, OTHMAN A S. Identification of differentially expressed genes and signaling pathways in bark ofseedlings associated with secondary laticifer differentiation using gene expression microarray[J]. Plant Physiology and Biochemistry, 2016, 107: 45-55.
[10] WANG X, SHI M, WANG D, CHEN Y, ZHANG S, WANG L, TONG Z, TIAN W. Comparative proteomics of primary and secondary lutoids reveals that chitinase and glucanase play a crucial combined role in rubber particle aggregation in[J]. Journal of Proteome Research, 2013, 12: 5146-5159.
[11] TANAKA Y, AIK-HWEE E, OHYA N, NISHIYAMA N, TANGPAKDEE J, KAWAHARA S. WITITSUWANNAKUL R. Initiation of rubber biosynthesis in: characterization of initiating species by structural analysis[J]. Phytochemistry, 1996, 41(6): 1501-1505.
[12] TANGPAKDEE J, TANAKA Y, WITITSUWANNAKUL R, CHAREONTHIPHAKORN N. Possible mechanisms controlling molecular weight of rubbers in[J]. Phytochemistry, 1996, 42(2): 353-355.
[13] BLACK L T, HAMERSTRAND G E, NAKAYAMA F S, RASNIK B A. Gravimetric analysis for determining the resin and rubber content of guayule[J]. Rubber Chemistry and Technology, 1983, 56: 367-371.
[14] SUCHAT S, PIOCH D, PALU S, TARDAN E, NICOLAAS E, DAVRIEUX F. Fast determination of the resin and rubber content inbiomass using near infrared spectroscopy[J]. Industrial Crops and Products, 2013, 45: 44-51.
[15] 姚行成, 陈先红, 周 珺, 王 军, 林位夫. 巴西橡胶树初生乳管中天然橡胶的提取及橡胶特性初步研究[J]. 热带农业科学, 2017, 37(11): 76-78.
YAO X C, CHEN X H, ZHOU J, WANG J, LIN W F. The primary study on extraction and characteristics of rubber from primary laticifers in[J]. Chinese Journal of Tropical Agriculture, 2017, 37(11): 76-78. (in Chinese)
[16] SALEHI M, CORNISH K, BAHMANKAR M, NAGHAVI M R. Natural rubber-producing sources, systems, and perspectives for breeding and biotechnology studies of[J]. Industrial Crops and Products, 2021, 170: 113667.
[17] HODGSON-KRATKY K J, STOFFYN O M, WOLYN D J. Recurrent selection for rubber yield in Russian dandelion[J]. Journal of American Society for Horticultural Science, 2017, 142: 470-475.
[18] SCHMIDT T, LENDERS M, HILLEBRAND A, DEENEN N, MUNT O, REICHELT R, ESENREICH W, FISCHER R, PRUFER D, GRONOVER C S. Characterization of rubber particles and rubber chain elongation in[J]. BMC Biochemistry, 2010, 11: 11.
[19] SAKDAPIPANICH J T, SUKSUJARITPORN S, TANAKA Y. Structural characterization of the small rubber particles in freshlatex[J]. Journal of Rubber Research, 1999, 2(2): 160-168.
[20] TARACHIWIN L, SAKDAPIPANICH J T, TANAKA Y. Relationship between particle size and molecular weight of rubber from[J]. Rubber Chemistry and Technology, 2005, 78: 694-704.
[21] BONFILS F, FLORI A, SAINTE-BEUVE J. Relations between wallace plasticity and Mw for natural rubber[J]. Journal of Applied Polymer Science, 1999, 74: 3078-3087.
[22] YIP E. Clonal Characterization of latex and rubber properties[J]. Journal of Natural Rubber Research, 1990, 5: 52-80.
[23] BONFILS F, DOUMBIA A, CHAR C, SAINTE-BEUVE J. Evolution in the natural rubber native structure and plasticity retention index from the first tapping of clonal trees[J]. Journal of Applied Polymer Science, 2005, 97: 903-909.
橡胶树初生乳管橡胶的产量性状与橡胶特性研究
姚行成1,2,涂寒奇1,2,周 珺1,2,陈先红1,2,陈 青1,2,林位夫1,2,王 军1,2*
1. 中国热带农业科学院橡胶研究所,海南海口 571101;2. 农业农村部橡胶树生物学与遗传资源利用重点实验室,海南海口 571101
巴西橡胶树()的乳管分初生乳管和次生乳管2种类型,前者分布在叶片和初生生长茎干的皮组织,后者分布在次生生长茎干的皮组织。目前,商用橡胶主要来自次生乳管,关于次生乳管橡胶的认识较透彻;由于初生乳管橡胶未被开发利用,因此人们对其了解也较少。以来自橡胶嫁接幼树枝叶的初生乳管橡胶为研究对象,通过分析与初生乳管橡胶产量相关的因子,包括枝叶生物量、生物量组成和橡胶含量等,明确初生乳管橡胶的产量潜力。同时,还研究了初生乳管橡胶的生胶性能和硫化特性。结果表明,橡胶幼树枝叶的生物量、生物量组成和橡胶含量因无性系而异,受无性系影响。在枝叶生物量组成中,叶片、树皮和木质杆分别占枝叶生物量的55.5%、10.5%和34.0%。叶片和树皮的橡胶含量分别为每克干物质含橡胶13.7和35.6 mg。初生乳管胶乳的橡胶粒子平均粒径(0.36 μm)和相对分子量低于常规割胶树的胶乳橡胶。初生乳管橡胶的氮含量和PRI分别为0.56%和89%,均高于常规橡胶的氮含量(0.50%)和PRI(80%)。初生乳管橡胶的门尼粘度和塑性初值0分别为66和26,均低于常规橡胶的门尼粘度(101)和0(50)。初生乳管橡胶的挥发份和灰分含量均与常规橡胶相同,分别为0.50%和0.38%。初生乳管橡胶的初始硫化时间10为2.00 min,正硫化时间90为19.12 min,均比常规橡胶的102.47和9025.08硫化时间短。初生乳管橡胶硫化曲线的最小扭矩较小(0.22 dN·m),而最大扭矩较大(8.25 dN·m)。通过分析3个无性系材料,结果表明萌条枝叶生物量、生物量组成和橡胶含量具有品系特征,并影响初生乳管橡胶产量,这为初生乳管橡胶在资源育种与栽培技术指明了研究方向。鉴于初生乳管橡胶的门尼粘度和0达不到标准胶要求,但其他理化性能指标与常规橡胶相似,建议初生乳管橡胶不能作为标准胶使用,但可作为普通胶料用于生产。
橡胶树;天然橡胶;初生乳管;理化性能
S794.1
A
2022-04-02;
2022-06-06
中国热带农业科学院基本科研业务费专项(No. 1630022015025);现代农业产业技术体系天然橡胶产业体系岗位科学家项目(No. CARS-33-YZ4)。
10.3969/j.issn.1000-2561.2022.11.004
姚行成(1980—),男,博士,助理研究员,研究方向:橡胶树栽培技术。*通信作者(corresponding author):王 军(WANG Jun),E-mail:wangjuncatas@163.com。
