阳春砂花丝、花柱转录组测序及生物信息学分析
2022-12-13杨博涵汤丽云李子翔何国振
杨博涵, 徐 杰,2, 汤丽云, 苏 景, 李子翔, 何国振
[1.广州中医药大学中药学院,广东广州 510006; 2.广东一方制药有限公司/广东省中药配方颗粒企业重点实验室,广东佛山 528244;3. 华南农业大学生命科学学院,广东广州 510642; 4.阳春市农业试验场(阳春市春砂仁试验场),广东阳春 529600]
阳春砂(Amomumvillosum)是姜科(Zingiberaceae)豆蔻属(Amomum)多年生常绿草本植物[1],其干燥成熟果实为四大南药之首——砂仁的主流品种,具有理气安胎、温脾止泻、化湿开胃的功效[2]。作为传统的大宗药材,阳春砂每年的需求数量大于300万kg,具有重要的药用价值及经济价值[3]。但阳春砂在生产上存在着严重的低产问题,其自然结实率仅有1.1%[4],研究者对此进行了许多研究,认为较低的自然结实率与阳春砂小花的特殊花器结构有着密切的联系。何国振等将阳春砂特殊的花器结构称为假合蕊柱,即雌雄蕊贴合在一起,但并未完全合生,在处于开花的状态下,唇瓣包裹着假合蕊柱,并且呈现出半抱合的状态,与唇瓣间的距离极近,仅有1.3~2.0 mm,严重阻碍了相应虫媒的授粉[3]。为提高产量,农户在实际种植中主要采用人工授粉的方式进行劳作,而人工授粉成本高、劳作强度大、根状茎及花序易被严重踩踏损伤等原因是传统人工授粉方式固有的弊端,严重抑制了农户的生产积极性。
细胞伸长或者细胞分裂所致的植物器官运动与内源激素及相关抑制剂作用相关,而细胞结构及生长速率的不对称是花器官运动的基础。有文献研究报道,腊梅(Chimonanthuspraecox)花丝的运动来源于其两侧表面细胞生长速率的差异[5]。姜科植物马来良姜(Alpiniamutica)花柱运动部位两侧细胞层数的差异是其花柱卷曲运动的基础[6],赤霉素(GA)、茉莉酸(JA)、吲哚-3-乙酸(IAA)等激素则有着调控植物雌雄蕊发育的作用[7-9]。何卓航等研究发现,随着阳春砂小花的生长,其花丝、花柱远、近轴侧的细胞层数出现了差异,这种不对称的结构是雌雄蕊运动的基础,认为阳春砂雌雄蕊的相向运动致使其假合蕊柱的形成[10];同时对花柱的近端以及远端轴侧的IAA水平进行测定,结果表明,生长时期在保持同样的水平不同横切部位中,IAA水平基本上呈现远端轴侧小于近端轴侧的趋势,故花柱运动的原因可能是来自于花柱近、远轴侧IAA水平的差异,该研究从生理层面初步阐述了阳春砂假合蕊柱的形成机制。然而,若要进一步探明阳春砂假合蕊柱形成机制,则须对阳春砂花器官运动及发育的分子机制作深入研究。
因此,本研究对阳春砂不同生长时期的花丝、花柱进行转录组测序,通过数据拼接、组装的方式建立阳春砂花丝、花柱的转录组数据库,将所得的Unigene进行功能注释、分类以及简单重复序列(SSR)分子标记。同时,重点关注并筛选可能影响阳春砂雌蕊、雄蕊运动的激素合成与信号转导途径关键基因。以期进一步解析阳春砂假合蕊柱产生的调控机制及分子机制,并为人为干预阳春砂的花器官结构,提高产量奠定理论基础。
1 材料与方法
1.1 试验材料
材料为大田栽培的不同生长时期阳春砂小花的花柱及花丝,阳春砂植株栽培于广东省阳春市合水镇那软村阳春砂种植基地(22°17′N,112°01′E),于2017年5—7月在该基地开展大田试验。参照陈红的方法[11],按照长度的不同,将阳春砂小花的生长时期进行了划分(表1),并于阳春砂花期内,每天上午采摘小花,解剖并分离出不同时期小花的花丝与花柱,迅速置于液氮灌中,随后转移至超低温冰箱(-80 ℃)进行保存。
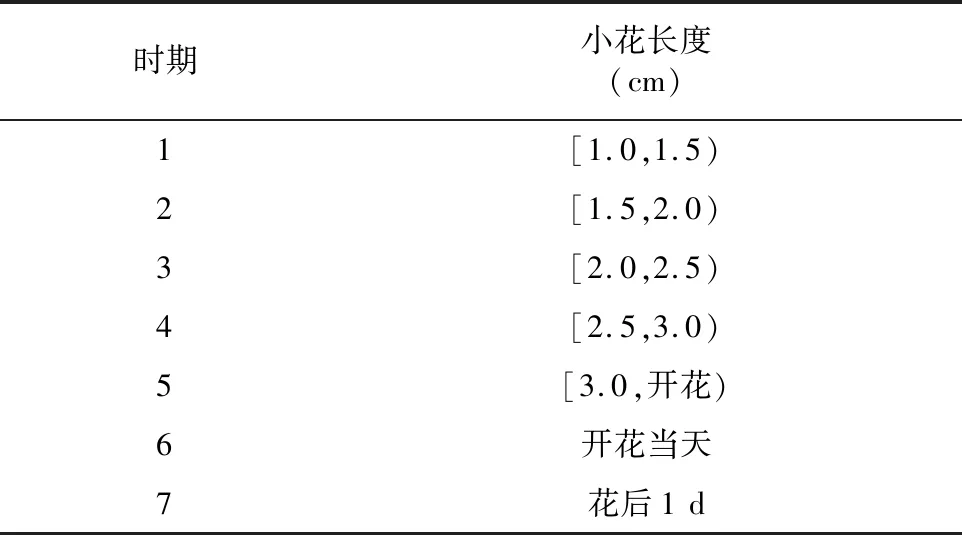
表1 阳春砂小花生长时期划分
1.2 试验方法
1.2.1 RNA提取 分别提取阳春砂小花在各生长时期的花柱和与花丝的总RNA,重复3次。利用NanoDrop 2000和Agilent 2100 Bioanalyzer测定RNA的浓度、纯度与完整性,数据检测合格后的RNA用于构建转录组数据库。此部分试验与转录组的测序委托深圳华大基因科技服务有限公司完成。
1.2.2 cDNA文库构建及转录组序列组装 使用Oligo(dT)磁珠富集质检合格且所有样品的混合RNA。加入打断试剂,以片段化的mRNA为模板合成1链、2链cDNA,配制反应体系,使接头与cDNA连接。PCR反应及产物回收、扩增。PCR产物变性,充分混匀,得到单链环形产物,随后PCR产物变性,即得到文库。利用Agilent 2100 Bioanalyzer和ABI StepOnePlus real-time PCR System对所得文库进行检测,检测合格后进行转录组denovo测序。所得reads通过Trinity软件进行序列组装,组装序列的质量通过BUSCO软件进行评估。
1.2.3 转录组基因功能注释及数据挖掘 利用生物信息学方法分析获得的阳春砂Unigene,为获得全方位的基因功能信息,对组装所得的Unigene在七大功能数据库中进行注释,包括NR、NT、 KOG/COG、GO、KEGG、SwissPro及Interpro。使用MIcroSAtellite (MISA)工具对Unigene进行SSR位点的挖掘。并在KEGG代谢通路中重点关注植物激素生物合成与信号转导Unigene的注释情况。
2 结果与分析
2.1 转录组数据组装
使用Illumina HiSeq-2000平台对不同时期的阳春砂花丝及花柱的cDNA进行测序。测序的结果显示,从转录组中获得了81.40 Mb的原始读数,用过滤软件SOAPnuke去除低质量的reads后共得到68.48 Mb Clean reads,最终获得10.33 Gb的碱基总数。Q20和Q30的百分比分别为97.20%和93.16%(表2)。说明转录组的测序质量较高,可以满足后续的生物信息学分析。
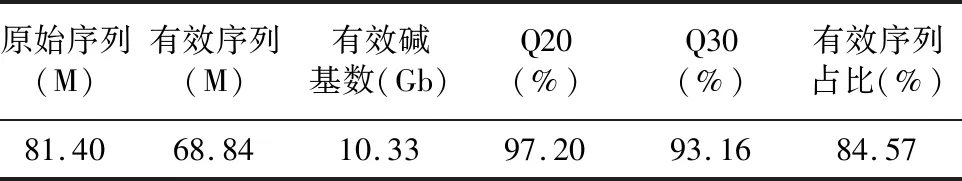
表2 测序数据质量分析
对转录本中Clean reads进行组装,一共获得138 590个转录本,包含了111 312 938个核苷酸的序列信息,这些片段长度的平均值为803 bp,N50为1 414 bp,GC含量为44.28%。对转录本进一步聚类并去除冗余的序列之后得到94 584条Unigene,总长度为 92 501 015 bp。Unigene的N50、N70和GC含量分别为 1 582bp、1 002 bp和44.35%,均大于其对应平均长度,说明组装的结果良好(表3)。

表3 转录本和单基因簇统计分析
2.2 阳春砂转录组基因总体注释情况
将所获得的结果在七大功能数据库进行注释,结果见表4。结果显示,阳春砂共注释到94 584条Unigene。其中,共有58 669条Unigene被NR数据库注释,占比最多,达到总Unigene的62.03%;NT数据库有40 205条,占42.51%;SwissProt有40 318条,占总Unigene的42.63%;KEGG数据库与KOG数据库分别有44 000条和45 892条Unigene,各占46.52%与48.52%;Interpro有45 793条,占48.42%。GO数据库注释到的基因最少,仅有 12 050 条,占总数的12.74%;所得比对结果显示,在七大数据库中均能成功注释的Unigene共有 5 691 条,占总Unigene条数的6.02%。
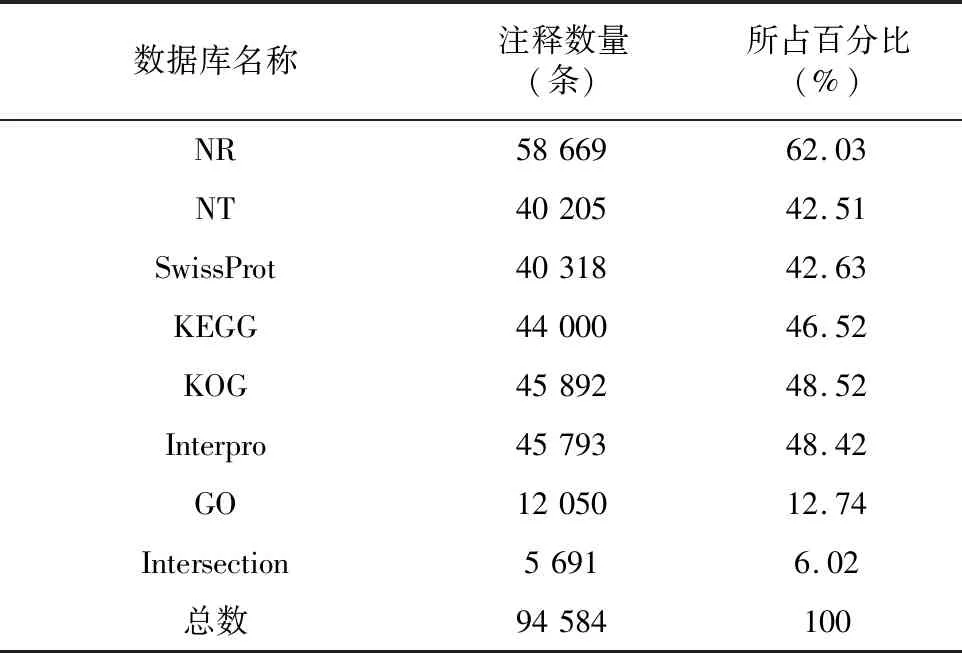
表4 转录组基因注释情况统计
2.2.1 NR数据库功能注释 NR数据库的注释结果(图1)显示,匹配最多的物种为小果野芭蕉(Musaacuminatasubsp.Malaccensis),该物种注释到的基因数量最多,共有45 257条,占比高达77.14%,证明阳春砂与该物种的同源性较高;其他物种依次为油棕(Elaeisguineensis)与海藻(Phoenixdactylifera),分别有3 338、2 435条Unigene被注释,占比分别为5.69%、4.15%,阳春砂与这2种植物的同源性相对较低。而剩余的13.02%则分布于其他物种中。

2.2.2 KOG 数据库功能注释 在KOG数据库中,共有45 892条Unigene被注释到(图2),共分为25个大类功能区,包括一般功能预测、信号转导机制、转录等功能。在为数众多且不同的功能分类中,注释到的基因的数量差异较为显著,一般功能预测类基因数量最多,共有11 177条Unigene,占比为24.36%,其次是信号转导机制,有8 271条Unigene被注释,占比为18.02%;除此之外,负责转录功能的Unigene有5 470个,占比为11.92%;有1 172条Unigene注释到负责次生代谢产物生物合成、运输和代谢功能区中,占比为2.55%。
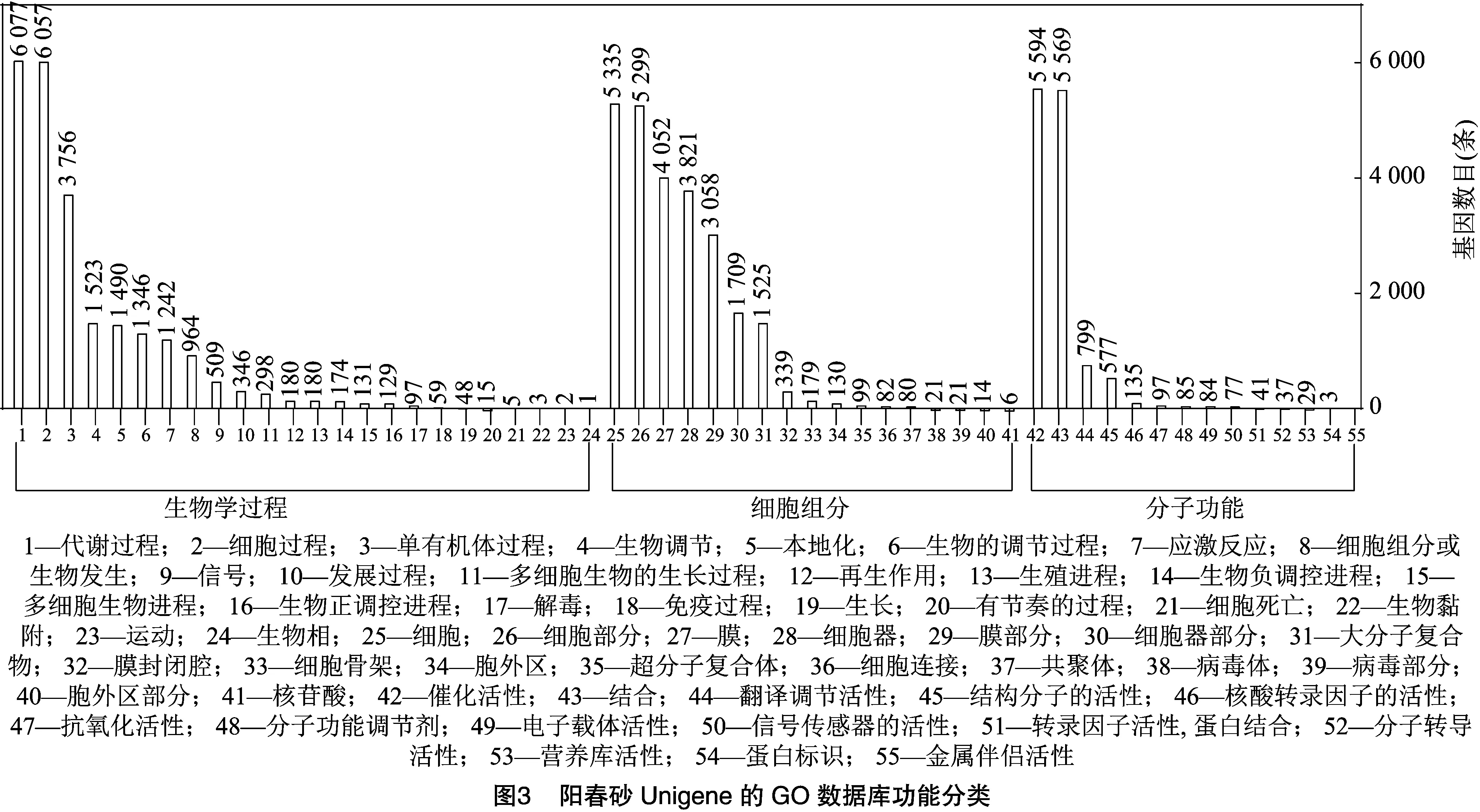
2.2.3 GO数据库功能注释 GO数据库中的功能分类注释结果见图3,结果表明,阳春砂花丝及花柱中共有12 050条Unigene注释到不同的功能节点上,共涉及到生物学过程、细胞组分和分子功能3个大类,55个亚类。归入到细胞组分的17个亚类中,以细胞、细胞部分、膜和细胞器功能的Unigene数量最多,分别有5 335、5 229、4 052、3 821条。在分子功能的14个亚类中,催化活性和结合注释数量最多,分别有5 594、5 569条。而涉及生物学过程的24个亚类中,以代谢过程(6 077条)和细胞进程 (6 057 条)为主。结果表明,同一个Unigene可以注释到多个功能结点上, 因此同一个功能分支的总注释数大于注释到该功能分支的总Unigene数。

2.2.4 KEGG数据库功能注释 对转录组测序获得的Unigene进行KEGG代谢通路富集分析后发现,共有44 000条 Unigene得到注释,涉及到137个通路。对富集显著的前20条通路进行分析(表5),其中,共有8 660条Unigene被代谢途径通路所注释,占比最多,达到注释总数的19.68%。而次生代谢产物的生物合成通路共有4 191条Unigene被注释到,占注释总数的9.53%,另外,本研究主要关注的植物激素转导通路一共有1 656条Unigene被注释到,占比为3.76%。
在所有的代谢通路中,共有23个与次生代谢相关的代谢通路,对其所有的Pathway进行分析后(表6)发现,一共有4 191个Unigene被次生代谢产物的生物合成途径所注释;而在本研究主要关注的植物激素合成途径中,62个Unigene被油菜素内酯生物合成途径所注释,另有59个Unigene被玉米素生物合成途径所注释。
2.2.5 SSR特征分析 转录组测序所得Unigene的SSR的检测最终结果显示,共有18 895个SSR被检测到(图4)。总共有14种多碱基重复SSR,其中, 三核苷酸重复的SSR 数目最多,共有6 313个,占SSR位点总数的33.41%;三核苷酸重复中AGG/CCT重复基元最多,共有1 434个;单核苷酸和二核苷酸重复分别有6 238个(33.01%)和5 128个(27.14%),二核苷酸中重复基元最多的是AG/CT,共有2 909个;六核苷酸重复数目共有546个(2.89%),四核苷酸重复330个(1.75%)和五核苷酸重复340个(1.80%)。
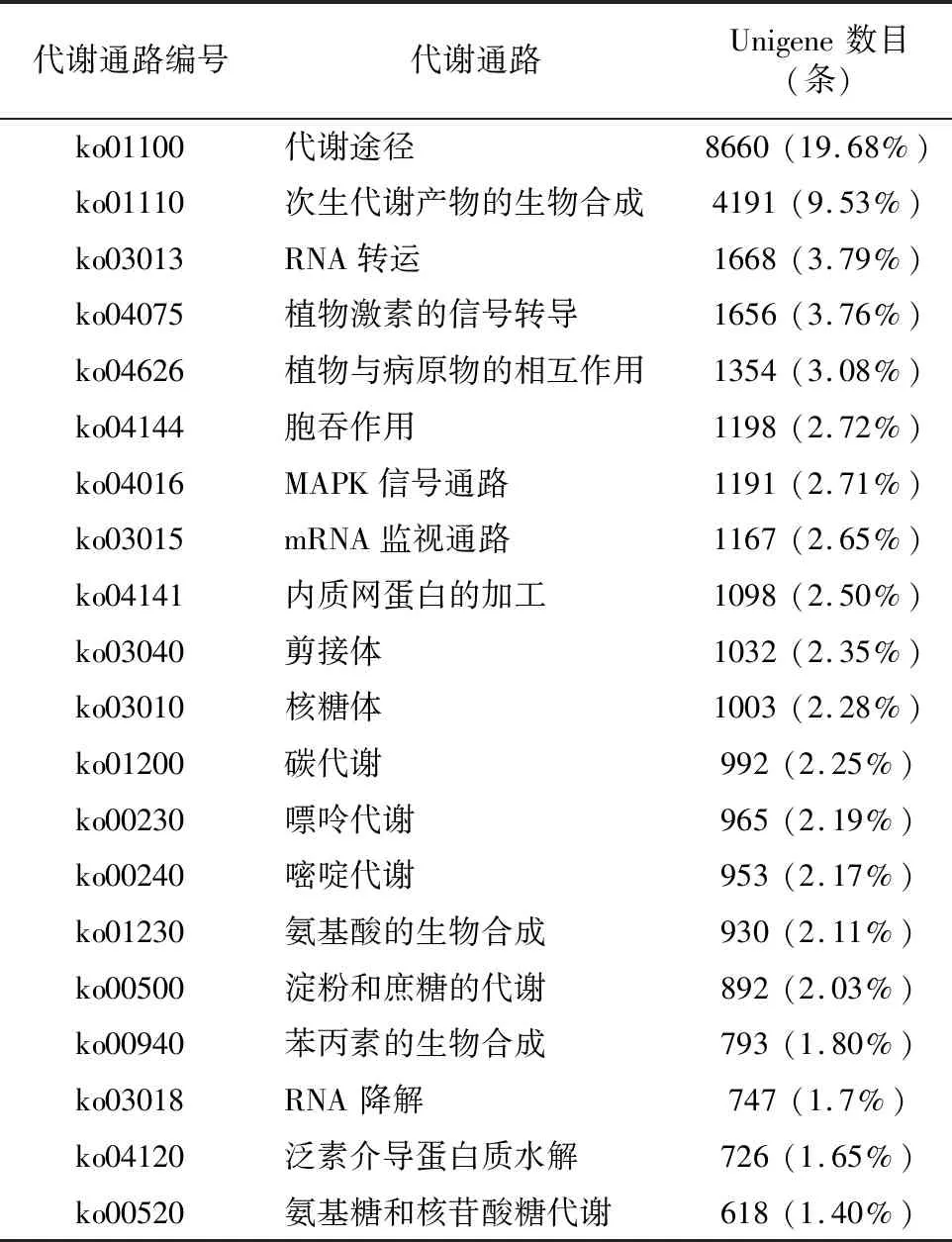
表5 次生代谢相关代谢通路的注释情况

表6 次生代谢相关代谢通路的注释情况

2.3 重点关注通路的Unigene 注释情况
2.3.1 植物激素信号转导途径中Unigene注释情况 在KEGG代谢通路中,共有1 760条Unigene注释到植物激素的信号转导途径中(表7)。赤霉素的信号转导途径中注释到的5个基因,对应的Unigene最多,共有438个;其中,植物光敏色素互作因子(PIF)对应的Unigene最多,达到318个;而水杨酸信号转导途径注释到3个基因,包括非表达病程相关蛋白(NPR1)、TGA转录因子及病程相关蛋白1(PR1)基因,但对应的Unigene最少,仅有74个;茉莉酸的信号转导途径中注释到的茉莉酸ZIM结构域蛋白(JAZ)、茉莉酸氨基酸合成酶(JAR1)、MYC2转录因子和茉莉酸受体蛋白(COI1)一共对应了176条Unigene;乙烯和油菜素内酯信号转导途径注释到的基因最多,均为8个,分别注释到139条及191条Unigene;其他植物激素中,生长素、细胞分裂素以及脱落酸的信号转导途径分别注释到329、245、168条Unigene;统计结果中,各激素信号转导途径关键基因均有注释,如细胞分裂素中的磷酸转移蛋白(AHP)、生长素中的生长素载体(AUX1)、生长素/吲哚乙酸蛋白(Aux/IAA)转录家族以及脱落酸信号转导途径中的ABF转录因子等。综上,研究较为深入的植物激素基本均在KEGG通路中有所注释,且其信号转导途径的关键基因都有一定数量的Unigene被注释到。

表7 转录组中植物激素信号转导途径Unigene注释情况
2.3.2 植物激素合成途径中Unigene注释情况 转录组中激素合成途径酶基因Unigene注释情况(表8)显示,赤霉素注释的酶基因最多,包括内根-贝壳杉烯合成酶(KS)、古巴焦磷酸合成酶(CPS)、内根-贝壳杉烯氧化酶(KO)及GA3氧化酶(GA3ox)等8个合成通路中的关键酶基因,一共对应67条Unigene;油菜素内酯与水杨酸合成途径中均有1个酶基因被注释到,为异分支酸合酶(ICS)与油菜素内酯-6-氧化酶2(CYP85A2),分别对应9条和24条Unigene;茉莉酸合成途径2个关键酶基因丙二烯氧化物合成酶(AOS)以及丙二烯氧化物环化酶(AOC)分别注释到6条和7条Unigene,生长素一共注释到42条Unigene,色氨酸转氨酶(TAA1)及吲哚-3-丙酮酸单加氧酶(YUC)分别注释到了其中22条和20条;乙烯合成途径中的乙烯合成前体合成酶(ACS)与乙烯合成前体氧化酶(ACO)分别对应9和14条Unigene;脱落酸及玉米素的生物合成途径中均注释到3个关键酶基因,分别有29条和45条Unigene。

表8 转录组中植物激素生物合成途径Unigene 注释情况
3 讨论与结论
近年来,转录组测序技术发展迅速,同时伴随着多个生物信息学分析平台的加入,大量研究即便在没有基因组数据支撑的前提下,通过转录组测序所得的结果也可在挖掘植物未知基因、明确相关生理功能的代谢途径及基因调控机制等方面提供海量相关信息。无需参考基因组的数据便可对目标物种进行分子生物学方面的研究,已成为中草药材基因信息挖掘的重要研究手段[12]。利用该技术,前人已从多种植物中挖掘出有关花器官发育的基因,并分析了相关基因的调控机制。李梅等利用雌蕊缺失茶树(Camelliasinensis)花的花、花蕾、花芽作为材料,基于转录组测序技术,共发掘出了34个花器官发育差异表达的相关基因,包括KNOX家族基因、WUS类基因、花器官发育 ABCDE 模型的相关基因等,证明茶树花即使是在雌蕊缺失的情况下,有关雌蕊发育的基因也是同样存在[13]。而在长瓣兜兰(Paphiopedilumdianthum)花蕾和花朵的转录组测序中发现,相较于花蕾期,花朵时期中调控花朵发育相关的基因,如AG、C2H2-ZEP等基因的表达量明显下调,大量基因表达下调的原因可能是在花朵时期,花的各部分器官已完成分化,调控器官分化基因的相关任务已完成[14]所致。位明明等通过该技术初步明确了多个参与编码橡胶树(Heveabrasiliensis)花器官发育的基因家族,如Mads-box、MYB及AP2等基因家族[15]。
本研究通过RNA-Seq技术构建了阳春砂不同时期花丝以及花柱总样品的转录组数据库,共获得94 584条Unigene,Unigene的平均长度为842 bp,N50为1 582 bp,GC含量为44.35%。其中,共有 62 174 条Unigene被注释到GO、KEGG、NR和KOG等七大数据库,占总Unigene的 65.73%。注释率较高,测序所得结果与拼接质量较好。这些注释的Unigene为阳春砂的代谢途径、基因功能分类、花器官发育及雌雄蕊运动分析等方面提供参考依据。此外,一共有32 410条Unigene未被注释到,占总Unigene的34.27%,推测这些Unigene可能为阳春砂中的非编码 RNA 序列或现有基因数据库尚未完善所致。在NR数据库中共注释到58 669条Unigene,其中77.14%的Unigene被注释到姜科植物小果野芭蕉中,两者比对所得同源基因数目较多的原因可能是亲缘关系较近。转录组所得Unigene在GO功能分类中,注释到生物学过程、细胞组分以及分子功能三大类,分别有 24、17、14个亚类,主要集中在代谢过程、催化活性、细胞部分和细胞过程等功能。而在KOG数据库的分析结果中,共有 45 892 条Unigeine得到了注释,在25个功能大类中,一般功能预测的Unigene最多,共有11 117条,信号转导机制、转录以及翻译后的修饰、蛋白质转换与代谢功能次之,核结构与细胞运动最少,分别仅有346条和75条Unigene。通过KEGG数据库分析发现,被注释到44 000条Unigene参与了23类共计137个KEGG代谢通路,这些通路主要集中在代谢途径、次生代谢产物的生物合成、RNA转运以及植物激素信号转导等,可以通过所注释的通路全面了解阳春砂花丝及花柱的代谢途径信息。同时,在阳春砂花丝、花柱转录组中一共挖掘到18 895个SSR位点,在6种不同的核苷酸重复类型中均有分布,证明本研究转录组中位点的类型丰富,分布密度较大,具有良好的多态性潜能;在二核苷酸和三核苷酸中,重复最多的基序是分别是AG/CT和AGG/CTT,该结果与王焕的研究结果[16]一致。
植物激素是植物通过自身代谢所产生的一些有机信号分子,可在低浓度下产生明显的生理效应,并在合成部位发挥功能[17-18]。而花器官的运动受到激素调节,Luo等研究发现,外源施加IAA后,会明显影响姜科植物花柱的弯曲程度[19]。有些植物激素也会调控姜科植物蓝猪耳二长雄蕊的翻转与伸长[20]。此外,其他植物激素同样也在雌雄蕊发育中起着重要的调控作用。JA可调控花药的开裂,JA生物合成通路的相关基因DAD1与OPR3在生长素感知缺陷的拟南芥突变体中的表达量提高,可提前使得花药产生开裂,说明此过程是一种负向调节,即生长素可以通过介导JA的生物合成来调节花药的发育[21-22]。在模式植物拟南芥的其他研究中,茉莉酸与赤霉素信号转导相关基因也在拟南芥雄蕊的发育过程中起到了重要的调控作用[8,23-24]。在本研究重点关注的植物激素合成与信号转导途径中一共有2012条Unigene被注释,囊括了赤霉素、生长素、茉莉酸及脱落酸等9种重要的植物激素,为进一步从分子层面阐述阳春砂花器官的发育及运动提供了大量候选基因。
笔者所在课题组于2012年提出通过生理手段改变阳春砂的花器结构,以适应更多种类的昆虫传粉而提高产量的研究思路[25]。近10年来,已经在阳春砂的花芽分化规律[26]、花器结构特征[3]、假合蕊柱的形成过程[10]、某些生殖生物学特性[27]、落果规律[28]等多方面进行了研究,明晰了阳春砂生产过程中由于特殊花器官结构导致传粉昆虫种类受限和低产的原因。因此,下一步的研究方向需对本研究中所获有关阳春砂花器官的发育及运动的关键候选基因进行更深层次的功能分析与验证,并对阳春砂假合蕊柱形成与相关基因表达模式的关系进行分析与探讨,不仅可为深入开展阳春砂花器官发育与运动相关基因的克隆及相互作用的模式分析提供数据基础,也为进一步深入揭示阳春砂假合蕊柱的分子调控机制提供了理论依据,以期形成人为干预的效果进而改变阳春砂花器官的形态,拓宽阳春砂唇瓣与假合蕊柱之间的距离,为阳春砂的授粉虫媒打造一个相对有利的授粉环境,从而提高自然结实率,达到增加农民收入的目的。
