纳米粒子的植物促生效应
2022-11-23韩云华米素娟石晓琪钟天航
韩云华,米素娟,石晓琪,钟天航
(草地农业生态系统国家重点实验室,兰州大学草地农业科技学院,甘肃 兰州 730020)
英文中“nano”一词来源于希腊语,意为“矮小的、侏儒”,该词在1959年第一次被物理学家费曼用作表述纳米概念[1]。纳米材料是指三维尺度空间中至少有一维处于纳米量级(1~100 nm)的材料,而纳米粒子指至少有二维处于1~100 nm的纳米材料[2-3]。与常规材料相比,纳米材料表现出许多特殊性质,如小尺度效应、电学效应、力学效应和催化性能,已广泛应用在能源、化工、医疗、电子等行业。纳米材料包括自然纳米粒子和工程纳米材料两类[3]。其中工程纳米材料分为碳纳米材料、无机纳米材料、有机纳米材料、有机无机杂合材料等[4]。
纳米材料对植物影响的研究起源于科学家对纳米材料广泛应用危害生态环境的担忧。随着纳米材料应用范围扩展,在生产、运输、利用等环节不可避免地排放到环境中。2005年,Wang等[5]第一次提出了纳米安全的概念。随后,为评估纳米材料释放可能产生的后果,许多纳米材料毒理效应的研究相继开展。大部分研究结果表明,纳米材料在浓度较高时可对植物生长产生抑制作用。在浓度较低情况下,反而表现出一定的促生效应,如浓度<20 ppm时表现出抗氧化剂活性,提高植物抗逆性[6]。
进一步研究发现,纳米材料可通过肥料效应、植物生长调节剂效应、纳米酶效应等多重方式影响植物生长发育,实现提高抗逆性,提高植物产量,促进植物代谢产物合成等的效果,这是纳米农业提出的基础[2,7-8]。本研究就近年来纳米材料对植物的促生作用及相关生物学过程进行论述,并指出潜在的挑战和未来研究的重点,以促进纳米技术在植物生产中的研究与应用。
1 纳米材料在植物体内吸收和运输
纳米材料尺度极小,可经根、叶等器官进入植物体内[4]。目前发现,碳纳米管[9-12]、量子点[13-17]、金属纳米粒子[18-20]、金属氧化物纳米材料[21-23]等均可被植物吸收,但纳米材料吸收和功能发挥与其化学性质(如大小、带电性、表面官能基团、包被物质)密切相关[24-26]。如多壁碳纳米管(multi-walled carbon nanotube,MWCNT)的带电性和大小直接影响其在玉米(Zea mays)和大豆(Glycine max)组织、细胞和细胞器内的分布[27]。
纳米材料在根部可通过质外体途径和共质体途径被吸收,叶片除了以上途径外,还可经气孔进入(图1)[28]。跨膜过程中,纳米材料可能通过载体蛋白、细胞水孔蛋白、离子通道和内吞作用进入植物细胞[29]。纳米金处理后,在美洲黑杨(Populus deltoides×nigra)根部胞间连丝中发现有纳米金存在,表明纳米材料在细胞间传递也可能是通过胞间连丝途径[30]。
纳米材料不仅可进入植物组织和细胞,还可在组织间运输(图1)。纳米CuO(20~40 nm)颗粒被玉米根吸收后,经木质部导管运输至叶,也可通过韧皮部筛管由叶运输至根[31],纳米CeO2也具有类似的运输规律[32]。MWCNT引发种子后,幼苗维管束中观察到了其存在[33],经根部吸收后,在花瓣处出现了积累[11]。利用激光共聚焦显微镜观测发现,碳纳米量子点(carbon dots,CDs)被根吸收后经维管束质外体途径运输至茎和叶[15]。这些发现为揭示纳米材料发挥生物学效应奠定了理论基础,但目前已有研究大部分以定性研究为主,受制于技术条件,还缺乏定量研究,对纳米材料的转运效率研究还不够深入。已有学者针对纳米材料的吸收和运输做出了详细的论述[4,26,28,34-38]。
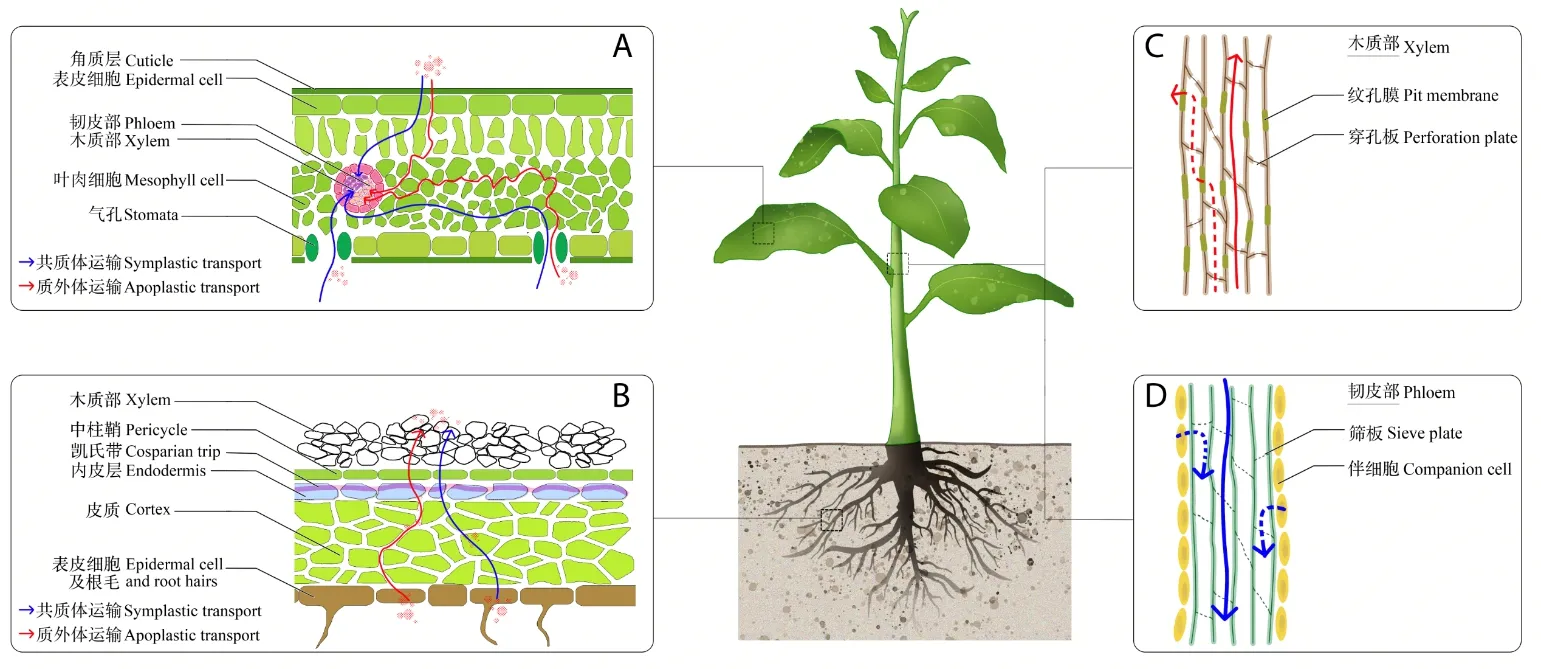
图1 纳米材料的植物吸收运输途径Fig.1 The absorption and transport pathways of nanomaterials in plants
2 纳米材料的营养效应
土壤养分是植物正常生长发育的基础。研究表明,近50%的作物产量获得与化肥使用有关[39]。传统肥料存在肥效短、效率低、易造成环境污染等缺点[40]。纳米材料的肥料效应主要表现在改变传统肥料的可利用性,如促进植物吸收、缓慢释放、降低环境风险等[41]。研究发现,尿素与羟基磷灰石纳米颗粒按6∶1混合后蒸干,形成的纳米复合物中,尿素释放速率为纯尿素的1/12,田间试验表明,纳米复合肥施氮量减少50%时,水稻(Oryza sativa)产量尚能稍高于施尿素处理,氮肥利用效率由施尿素的18%提高至48%[42]。值得注意的是,上述研究是在缺磷土壤中开展的,纳米羟基磷灰石也可同时提供磷元素和钙元素,对缺磷土壤中作物增产具有多重肥料效应。在国内也有类似研究,氮素减少20%,同时添加0.3%的纳米碳,N2O排放降低13.9%,小麦(Triticum aestivum)产量提高17.9%[43]。也有利用壳聚糖等纳米材料将传统肥料制成纳米复合物,在温室条件下,施用10~100 mg·L-1时,能显著缩短小麦生育期,增产51%~56%[44]。
除与常规肥料形成复合肥外,一些纳米颗粒亦可提供营养元素,促进植物增产。相比于常规CaO和Ca(NO3)2,叶面喷施纳米CaO提高了植株中Ca的积累量,促进根系的发育[45]。与MnSO4相比,叶面喷施0.05 mg·L-1纳米Mn促进了绿豆(Vigna radiata)根和茎的发育,生物量增加38%[46]。Fe2+与CDs形成的复合物缓解了甜瓜(Cucumis melo)缺铁症,提高了Fe2+转移效率[47]。目前已发现纳米Fe2O3、纳米Mg、纳米Fe、纳米CaCO3、纳米S、纳米Mn、纳米ZnO等都具有为植物提供营养的能力[48-49],但纳米肥料与常规肥料肥效差异研究还比较少[41,50]。
3 纳米材料的植物生长调节剂效应
植物生长调节剂是指天然的或人工合成的,低浓度即可影响植物发育和代谢过程的有机物[51]。前文论述了纳米材料的剂量效应,本研究将这种现象总结为纳米材料的植物生长调节剂效应。许多纳米材料均具有植物生长调节剂的效果,如前文论述的金属基纳米颗粒,但本研究将其归入纳米肥料或纳米酶的范畴。本部分主要讨论一类通常认为是植物生长非必需元素材料——碳基纳米材料。碳基纳米材料是纳米材料中重要的一类,包括碳纳米管(carbon nanotube,CNT)、CDs、石墨烯、富勒烯、碳纳米角(carbon nanohorns,CNH)等[52]。这类材料以碳为主要构成元素,具有生物毒性小、可降解的特点[53-54],亦可调控植物生长。研究表明,单壁碳纳米管(singlewalled carbon nanotubes,SWCNT)可提高小麦发芽率[55]。浓度0.1 mg·L-1的单壁碳纳米角(single-walled carbon nanohorn,SWCNH)可促进拟南芥(Arabidopsis thaliana)主根发育和侧根形成,基因表达分析表明与根系发育有关的基因YUC3、YUC5和PIN2表达量显著上调,代谢分析发现SWCNH重构了植物体内碳氮代谢[56];碳纳米管还可促进百脉根(Lotus corniculatus)、大豆等豆科植物根系结瘤[57-58]。此外,CDs、石墨烯、富勒烯等在豌豆(Pisum sativum)、菠菜(Spinacia oleracea)中均表现出促进种子萌发、根系发育、提高鲜重等效果[24,53,59-63]。
碳纳米材料提高植物抗逆能力。水培液中添加10 mg·L-1的MWCNT,改变了根系膜结构组成,提高水孔蛋白基因表达量,改善盐胁迫条件下根系吸水能力,增强了西兰花(Brassica oleracea)耐盐性[64]。水培液中添加CDs(180 mg·L-1)可促进根系生长,提高根系活力,增强花生(Arachis hypogaea)抗旱性,鲜干重、根长和苗长均为对照的1.5倍左右[65],SWCNT也可缓解植物轻中度干旱胁迫[66]。叶面喷施富勒烯改变了黄瓜(Cucumis sativus)代谢过程,提高了抗Cu胁迫的能力[67]。CDs可吸收Cd2+,并提高谷胱甘肽合成基因表达量,显著降低小麦和柚子树(Citrus maxima)根和叶片中Cd2+含量[68],促进拟南芥、柚子树根系发育,提高发芽率、生物量[69-70]。MWCNT通过表面吸附,亦可降低百草枯对植物的毒性[71]。
碳纳米材料调控植物生长效果与自身材料特性密切相关。Zhang等[72]利用L-和D-半胱氨酸合成具有手性的L-CDs和D-CDs,并处理绿豆后发现,两种类型CDs均可促进绿豆种子萌发、根系生长和地上部分生长,并提高根系活力和核酮糖-1,5-二磷酸羧化酶(Rubisco)活性,但D-CDs对根系活力和Rubisco活性的影响显著高于LCDs(28.9%和67.5% vs 8.4%和20.5%)。对鹰嘴豆(Cicer arietinum)的研究表明,直径小于10 nm的SWCNT要比直径约180 nm的MWCNT促生效果好,而直径约为4.5 nm的CDs促生效果较差[73]。
4 纳米酶与植物抗逆
一些纳米材料作为非生物酶,在提高植物抗逆性方面具有巨大的潜力。50 mg·L-1纳米CeO2颗粒显著提高香青兰(Dracocephalum moldavica)抗盐性[74]。浓度为100 mg·L-1的纳米ZnO可显著缓解玉米干旱胁迫,地上部干物质增加19.7%[75]。目前报道纳米铁、纳米ZnO、纳米TiO2、纳米CeO2、纳米SiO2、纳米Mn3O4、多羟基富勒烯等可提高植物抗旱性[76-79]、抗盐性[80-83]、抗重金属胁迫[84]等。
植物受到非生物胁迫时,会产生大量活性氧(reactive oxygen species,ROS),导致蛋白质、脂类、糖类和DNA氧化,清除ROS可显著提高植物抗逆性(图2A)[85]。Wu等[81]研究发现,CeO2纳米颗粒可清除叶片中ROS,但与其材料特征密切相关。丙烯酸包裹的CeO2颗粒(Ce3+/Ce4+=35.0%,粒径10.3 nm,电位-16.9 mV)使拟南芥叶片中ROS降低52%,但氨化的CeO2颗粒(Ce3+/Ce4+=60.8%,粒径12.6 nm,电位9.7 mV)增加了叶片中ROS含量。金属基纳米材料中,纳米Fe2O3、纳米ZnO、纳米TiO2、纳米Mn3O4也具有清除ROS的能力[86-87]。此类材料清除ROS是依靠氧化物中金属离子价态变化,如CeO2的催化机理如下[81]:
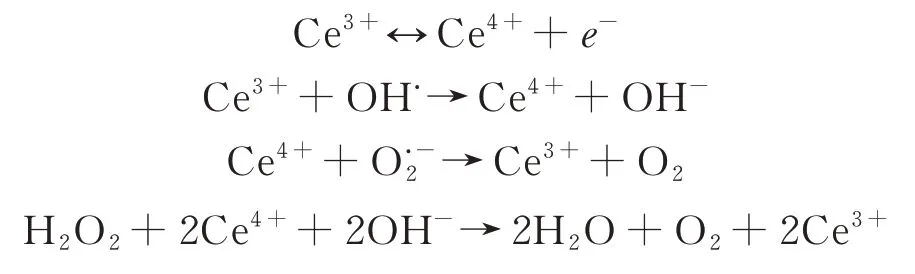
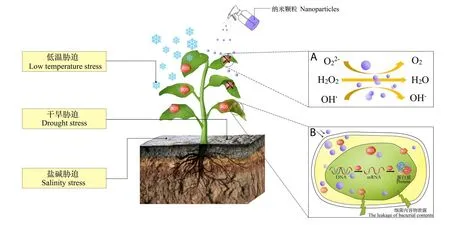
图2 纳米酶提高植物的抗逆机制Fig.2 Improvement of plant resistance mechanism by nano-enzyme
此外,富勒烯可定位于线粒体内,具有类超氧化物歧化酶(superoxide dismutase,SOD)效应,可显著提高植物抗逆性[77,88]。纳米材料与其他植物活性成分结合形成纳米复合物,也是一种提高植物抗逆性的新途径。研究发现,氮杂CDs(N-CDs)与辣根过氧化物酶(horseradish peroxidase,HRP)形成复合物N-CDs@HRP,可显著提高HRP活性,进而提高植物抗逆性[89]。
在影响生物胁迫方面,纳米材料具有优良的抑制病原微生物繁殖能力,降低植物病害程度。对禾谷镰孢菌(Fusarium graminearum)和梨孢镰刀菌(Fusarium poae)的研究发现,部分碳纳米材料在孢子表面聚集,影响孢子吸水,造成质壁分离是碳纳米材料可能的抑菌机理之一。单壁碳纳米管、石墨烯、氧化石墨烯均具有一定的抗真菌病原能力,其中单壁碳纳米管效果最好,500 μg·mL-1单壁纳米管使禾谷镰孢菌孢子萌发率降低95.2%[90]。此外,纳米TiO2、纳米ZnO、铜基、银基和壳聚糖纳米颗粒同样具有优异的广谱抗菌能力[91-93],相关机制见图2B。纳米材料亦可作为传统杀菌剂、杀虫剂的助剂,增加其防效,但这不是本研究讨论的重点,感兴趣可阅读Hajji-Hedfi等[93]的研究。
5 纳米材料与光合作用
光合作用通过光反应和暗反应两个阶段将空气中CO2合成有机物。纳米材料可影响光反应和暗反应的多个关键生化过程,提高植物光合速率[24,94]。光反应中,植物通过叶绿素吸收光能,光解水放出氧气(O2),产生高能电子(e-)和质子(H+),高能电子经一系列电子传递体的传递,还原NADP+为NADPH;同时,跨类囊体膜质子梯度(ΔpH)可驱动ATP合酶合成ATP[95]。纳米材料可影响植物叶绿素含量、光谱吸收、光系统(photosystem,PS)活性等,提高光反应速率。
叶绿素是叶绿体中主要的光吸收色素,其含量变化直接影响光反应捕获光的能力。研究发现土壤添加纳米TiO2(750 mg·kg-1)可提高黄瓜叶绿素含量[96]。氮杂CDs处理的绿豆叶绿素含量与对照相比提高14.8%,PSⅠ速率提高10.4%,碳水化合物量提高21.9%[61]。此外,纳米Zn、纳米Cu、纳米ZnO、纳米Fe2O3和纳米CeO2均可提 高植 物 叶 片 中 叶 绿素 含 量[68,97]。利 用qRT-PCR测 定 发 现,Mg,N-CDs处 理 上 调 了 叶 绿 素 合 成 基 因ChlI、ChlD、ChlG和chlorophyllase-2表达量[98]。
太阳光中,波长400~700 nm范围内的红蓝光可直接被叶绿体吸收,波长小于400 nm的紫外光不能被叶绿体直接利用。部分纳米材料可被紫外光激发,放出红蓝光,间接扩展了叶绿体吸收光谱[99]。将具有紫外-蓝光转换功能的CDs注射到烟草(Nicotiana tabacum)叶片中,在6 W紫外灯照射条件下,烟草叶片光合速率提高了18%[100]。纳米TiO2、CdSe量子点、单壁碳纳米管等均可实现紫外-红蓝光转换的功能,增加植物光合速率[101-102]。
纳米材料可通过影响PSⅠ和PSⅡ系统活性,提高光系统反应速率。纳米TiO2可提高希尔反应速率,增加氧气释放[103]。纳米Mn通过提高PSⅡ系统Mn4Ca复合物中的CP43蛋白活性,加速了水的光解和光合磷酸化效率[46]。有些纳米材料为良好的导体,可加速光反应中电子传递速率[82,104],如单壁纳米管可穿透叶绿体膜,在叶绿体质膜上积累,促进电子转移,光合作用效率提高3倍[105]。
暗反应阶段是利用光反应产生的NADPH和ATP,通过卡尔文循环固定CO2的过程[95]。此阶段纳米材料可调节气孔导度和关键限速酶活性,影响暗反应效率[94]。拟南芥中的研究发现,纳米Fe上调质膜H+-ATPase合成基因AHA2表达量5倍,促进了气孔的开放[106]。Rubisco是暗反应关键限速酶之一[107],可降解的CDs(5 nm,0.56 mg·mL-1)提高拟南芥Rubisco活性42%,种子产量增加20%[53]。有关纳米材料与光合作用关系的专题论述,可参阅Liu等[94]的研究。
6 研究展望
纳米材料在提高植物生产性能和抗逆性等方面具有广阔的应用潜力,但其种类多,材料特性多变,对植物的调控作用也因物种而异。广泛筛选易被植物吸收、促生效率高的纳米材料,并从生理、分子、细胞等角度研究植物对纳米材料的响应,仍是纳米材料调控植物生长研究的主要内容。
纳米材料调控植物生长的研究层出不穷,但目前大部分研究以植物生长初期为主,全生育期研究还比较少。此外,田间条件下的植物生产受到光、温、水、热、病虫害、土壤等多重环境因子影响,环境因素远比实验室内复杂,纳米材料在田间条件下促生效应研究应该给予更多的重视。
许多金属纳米材料包含重金属或稀土元素,如Mn、Cu、Ce、Ti等,虽然已有研究证实此类纳米材料对植物没有毒性或毒性很小,但此类研究都是基于植物生长周期特定阶段或一个植物生长周期。植物生产过程中势必要长期不断使用,因此引起的纳米材料环境累积效应还需要长期定位研究。此外,食物链具有顶端富集效应[96],有研究表明,纳米材料用于植物生产后,可在种子部分积累,并影响子代生长[108]。纳米材料在整个食物链的传递、富集效应还未受到广泛关注,开发可降解、环境友好的纳米材料是促进纳米材料应用的基础。
尽管面临众多挑战,纳米技术在植物生产中已经表现出巨大的应用潜力,未来的研究应采用系统性思维,从植物生产、环境保护、食物安全等角度全方位开展研究和评价,实现纳米技术的安全、高效利用,促进植物生产质量提升。
