灌浆期小麦旗叶在高温胁迫下的转录组分析
2022-11-17刘秀坤韩冉李晓明解树斌李法计陈亚鹏翟胜男李豪圣刘建军赵振东张玉梅曹新有
刘秀坤,韩冉,李晓明,解树斌,李法计,陈亚鹏,翟胜男,李豪圣,刘建军,赵振东,张玉梅,曹新有
(1.青岛农业大学农学院,山东 青岛 266109;2.山东省农业科学院作物研究所/小麦玉米国家工程中心/农业部黄淮北部小麦生物学与遗传育种重点实验室/山东省小麦技术创新中心,山东 济南 250100;3.山东鲁研农业良种有限公司,山东 济南 250100;4.滕州市农业农村局,山东 滕州 277599)
小麦是我国第二大粮食作物[1],2010—2019年平均种植面积在2400万公顷左右。小麦具有较高的营养价值和特殊的面筋特性,可用于多种食品的加工,为人类提供21%的食物热量和20%的蛋白质[2]。
小麦是喜凉作物,日均温20~24℃最适合其籽粒灌浆,超过30℃会对籽粒灌浆和品质造成影响[3]。研究表明,灌浆期高温天气频发会使小麦灌浆速率降低[4],灌浆时间缩短[5],造成小麦籽粒灌浆不充分,降低小麦产量;全球气温每上升1℃,小麦减产6%[6]。根据济南市气象局的温度记录,2017—2021年间小麦灌浆期日均温在27~30℃,最高可达35℃,这不仅会降低小麦产量,还会对小麦品质造成影响[7]。麦谷蛋白含量与醇溶蛋白含量的比值影响小麦的面筋特性,灌浆中后期高温,麦谷蛋白含量降低,醇溶蛋白含量增加,使麦谷蛋白含量与醇溶蛋白含量比值减小,降低面筋强度[8]。与此同时,灌浆期温度超过30℃,会使小麦淀粉含量降低,面团强度下降,影响其加工品质[9]。
植物在生长发育过程中往往会遭受诸多非生物因素胁迫,如盐碱、低温、干旱、高温等,这些非生物胁迫会使代谢过程紊乱,致使细胞内活性氧(ROS)积累,影响植物正常的生长发育[10]。目前,植物抵御高温胁迫的机理研究已取得了较大进展,与耐热相关的基因也在陆续挖掘和验证。田雪军在小麦中发现了2个耐热基因——TaMBF1c和TaMYB85,热胁迫条件下,TaMBF1c的缺失突变体在微管运输、胁迫响应、蛋白质加工代谢等途径中相关基因的翻译效率降低,而过表达TaMBY85基因的拟南芥和小麦耐热性提高[11]。高温胁迫会促进植物体热激蛋白(heat shock protein)的转录和翻译,而热激蛋白具有维持细胞结构稳定和正常生理功能的作用。Montero-Barrientos等研究表明,转哈氏芽孢杆菌hsp70基因的拟南芥表现出了更高的耐热性,且植株生长并没有受到抑制,在经过高温预处理后表现出了更好的抵御逆境胁迫的能力[12]。Katiyar-Agarwal等将拟南芥基因Athsp101导入水稻中,转基因水稻表现出更高的耐热能力,高温胁迫后产量没有显著下降[13]。在受到热胁迫后,小麦热激转录因子TaHsfA1能促进热激蛋白的表达,提高拟南芥幼苗的耐热性[14]。因此,明晰耐热调控网络,挖掘耐热相关基因,利用分子育种等手段选育耐热品种,对提高小麦耐热性具有重要意义。
本试验以本课题组前期研究中筛选到的小麦耐热品种菏麦13和热敏感品种临麦2号[15]为试材,经热胁迫处理,运用RNA-seq技术对灌浆期旗叶进行转录组分析,以挖掘热胁迫相关基因,为后续进一步研究耐热胁迫调控网络、选育耐热新品种奠定基础。
1 材料与方法
1.1 试验材料
本研究选用耐热品种菏麦13和热敏感品种临麦2号为材料,于2018年10月中旬正常播种于埋置在大田的花盆中,每盆10粒,按大田常规管理,待出苗后间苗,每盆留5株。每份材料种4盆。
1.2 样品处理及分类
花后14 d,将4盆材料(2盆菏麦13和2盆临麦2号)移至人工气候箱,42℃处理0、1 h,剩余4盆留至原处作为对照。取主穗同等部位的旗叶于-80℃冰箱保存。所取样品按照以下方案进行分类,用于后续的RNA-seq及分析。

表1 样品分类及差异分析对比方案
1.3 文库构建及测序
提取样品总RNA并使用DNaseⅠ消化DNA后,用带有Oligo(dT)的磁珠富集真核生物mRNA,向得到的mRNA中加入适量打断试剂,高温条件下使其片断化,再以片断后的mRNA为模板,合成cDNA,经过磁珠纯化、末端修复、3′末端加碱基A、加测序接头后,进行PCR扩增,从而完成整个文库制备工作。构建好的文库用Agilent 2100 Bioanalyzer和ABI StepOnePlus Real-Time PCR System进行质量和产量检测,文库质控合格后进行测序。
1.4 原始数据过滤统计及质量分析
应用base calling将测序得到的数据转化成序列信息,为了保证信息分析的可靠性,需对数据进行一系列的处理:去除含adapter的reads,去除含N比例大于10%的reads,去除低质量的reads(从整体上,Q≤20的碱基比例较低表明测序质量较好),从而获得clean reads。
1.5 差异基因的筛选
基于泊松分布分析方法,通过控制FDR(错误发生率),根据表达量(FPKM值)计算基因在不同样本间的差异表达倍数,以FDR≤0.001、≥2为标准筛选两样本间的差异基因。
1.6 转录表达谱qRT-PCR验证
为了验证转录组数据的可靠性,需要对差异表达基因进行荧光定量PCR验证。我们随机选取了20个差异表达基因进行qRT-PCR验证。利用Primer Premier 5软件进行引物设计(表2)。

表2 qRT-PCR引物
2 结果与分析
2.1 测序质量分析
从整体上看,样本的CG含量在53.34%~55.08%之间,Q20比例在98.95%以上,Q30比例在96.26%以上(表3)。说明样本的质量较好,可用于下一步分析。

表3 转录组测序数据统计
2.2 差异基因的筛选结果
最终4个对比方案共获得4715个差异基因(图1)。临麦2号叶片0 h-vs-临麦2号叶片1 h检测出1527个差异表达基因,菏麦13叶片0 h-vs-菏麦13叶片1 h检测出1190个差异表达基因,说明小麦旗叶中某些基因对高温胁迫做出响应,在高温条件下表达。临麦2号叶片0 h-vs-菏麦13叶片0 h检测到1225个差异表达基因,临麦2号叶片1 hvs-菏麦13叶片1 h检测到773个差异表达基因,说明在高温胁迫下某些基因会沉默,不表达。
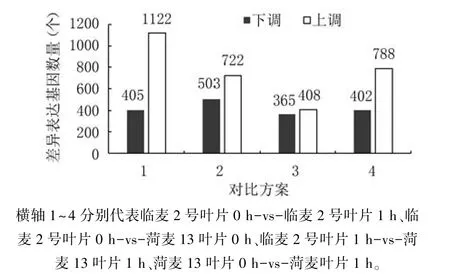
图1 样品差异表达基因统计
2.3 转录表达谱qRT-PCR验证
随机选择20个耐热基因进行qRT-PCR验证(图2),其中11个上调表达,如与逆境胁迫相关基因Rps7和Snf1、热胁迫相关基因MADS等;9个下调表达,如氧化胁迫相关基因Beta-carotene hydroxylase、核小体蛋白亚基H2B等。qRT-PCR验证结果表明表达趋势与RNA-seq测序分析结果一致,证明了RNA-seq测序结果的可靠性和可重复性,也为下一步确定候选基因并进行深入研究提供了参考。

图2 差异基因的qRT-PCR验证
2.4 差异基因GO显著富集分析
将获得的目的基因进行GO富集分析,主要从基因的分子功能(molecular function)、所处的细胞位置(cellular component)、参与的生物过程(biological process)三部分分析差异表达基因的生物学功能(图3)。4个对比方案中,生物过程主要富集在代谢过程、细胞过程、单体过程、定位和刺激反应,细胞组分部分主要富集在细胞、细胞组分、细胞器、细胞器组分、膜、膜组分和高分子复合物,分子功能部分主要富集在结合物和催化活性。
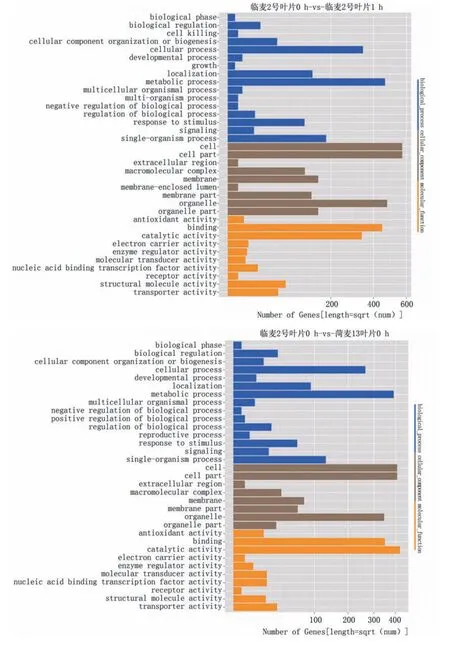
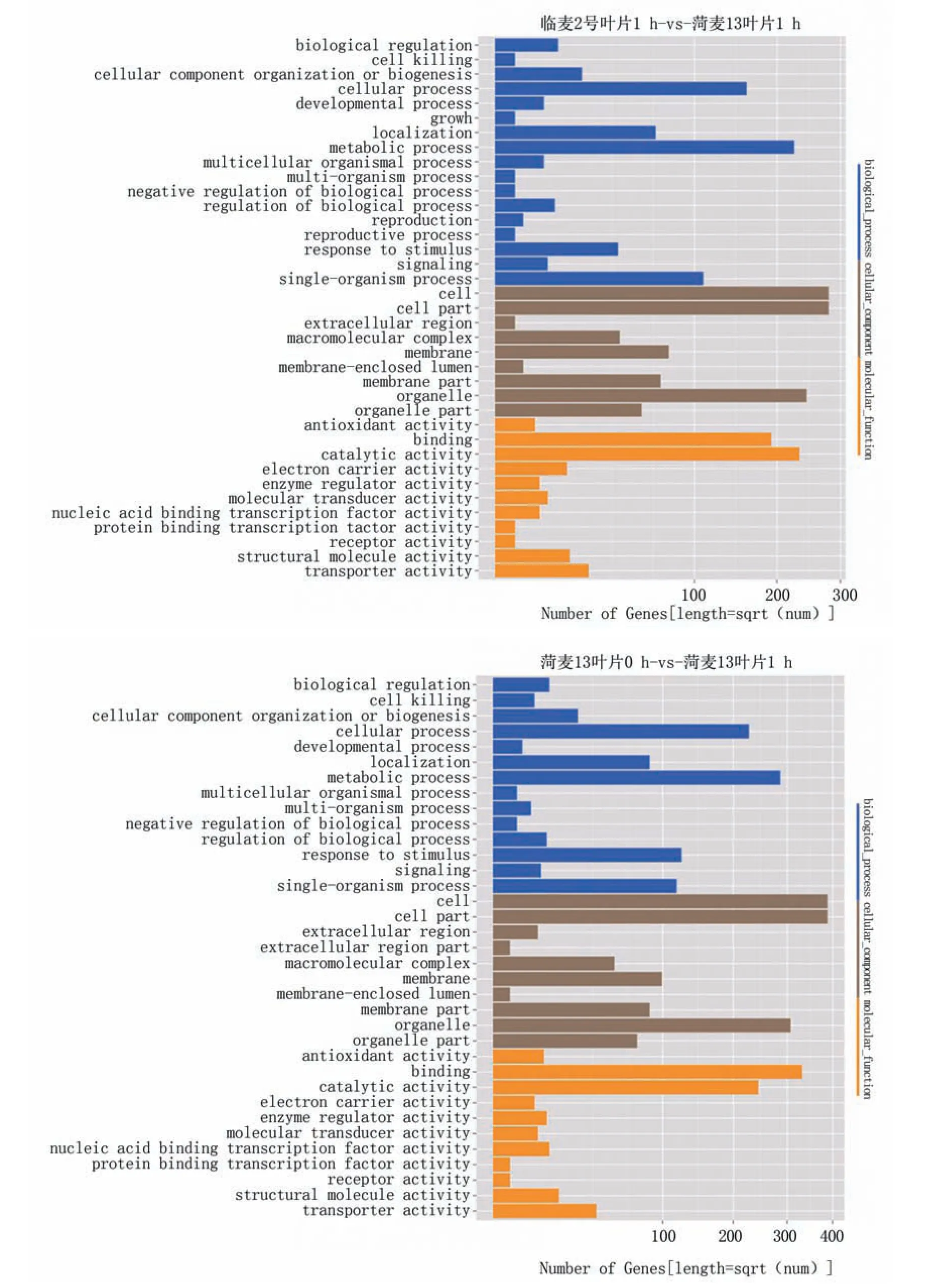
图3 差异基因GO功能富集分析
2.5 差异表达基因的KEGG注释及通路分析
通过对差异表达基因的通路注释可以进一步解释基因的生物学功能,对4715个差异表达基因进行KEGG分析,选取显著富集的前20个通路绘制散点图(图4)。临麦2号叶片0 h-vs-临麦2号叶片1 h之间差异显著富集的通路主要在内质网蛋白质加工、氧化磷酸化、内吞作用、剪接体、核糖体、光合作用、氮代谢、谷胱甘肽代谢,临麦2号叶片0 h-vs-菏麦13叶片0 h之间差异显著富集的通路主要在谷胱甘肽代谢、苯并恶嗪酮类化合物生物合成、二羧酸代谢、柠檬烯和蒎烯降解、维生素B6代谢、油菜素内酯的生物合成、二苯酚、二甲苯和姜二醇生物合成、次生代谢产物的生物合成、异黄酮的生物合成、亚麻酸代谢、甘油磷脂代谢、苯丙素的生物合成、自然杀伤细胞介导的细胞毒性、色氨酸代谢,临麦2号叶片1 h-vs-菏麦13叶片1 h之间显著富集的通路主要在代谢途径、光合作用、氧化磷酸化、二羧酸代谢、谷胱甘肽代谢、亚麻酸代谢、油菜素内酯的生物合成,菏麦13叶片0 h-vs-菏麦13叶片1 h之间差异显著富集的通路主要在内质网蛋白质加工、内吞作用、剪接体、异黄酮生物合成、光合作用、氧化磷酸化、柠檬烯和蒎烯降解、二萜生物合成、核苷酸切除修复、亚油酸代谢、错配修复、DNA复制。
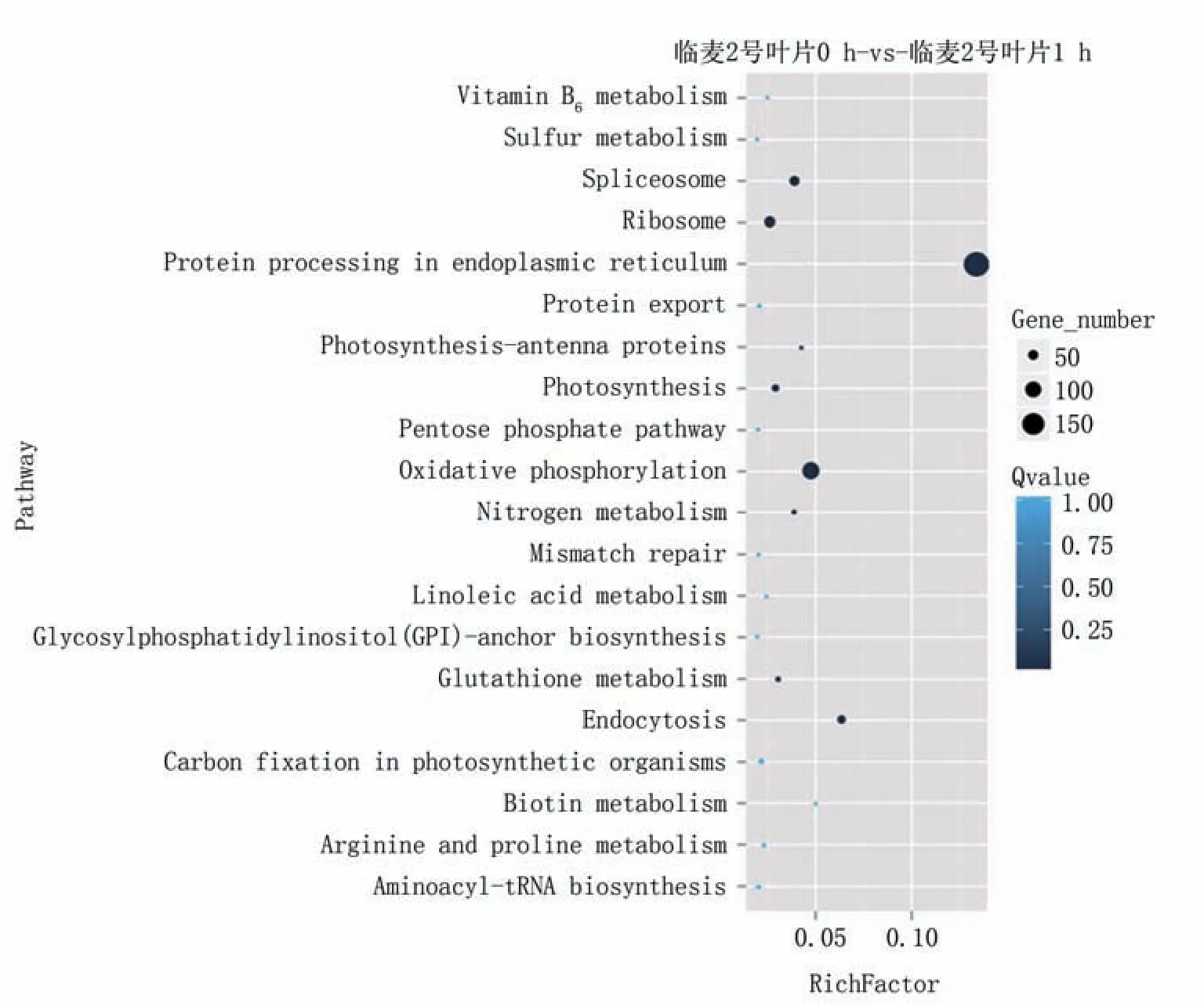


图4 差异基因KEGG富集通路
2.6 差异表达基因的转录因子
对所有差异表达基因归类,发现转录因子共有86个(表4),属于21个转录因子家族,主要包含AP2-EREBP家族15个(上调10个,下调5个),MYB家族11个(上调4个,下调7个),NAC家族8个(上调4个,下调4个),WRKY家族12个(上调5个,下调7个),HSF家族7个(上调7个)。

表4 差异表达基因转录因子数目统计
3 讨论与结论
植物对高温的响应是多基因控制的过程,为了明确小麦在全基因组水平上对高温的响应机制,本研究采用高通量测序技术,对耐高温小麦品种菏麦13和热敏感品种临麦2号在花后14 d 42℃高温处理0、1 h,取旗叶进行转录组测序分析,Q30比例在96%以上,表明测序质量良好,保证了数据的可靠性,可以进行后续分析。本研究共筛选出4715个差异表达基因,其中上调表达基因数目为3040个,下调表达基因数目为1675个。
3.1 不同品种在热胁迫下差异基因的GO分析
通过差异表达基因的GO富集分析可知,4个比较方案在参与的生物过程和所处细胞位置方面主要富集在代谢过程和细胞。在高温处理1 h后,临麦2号叶片0 h-vs-临麦2号叶片1 h、菏麦13叶片0 h-vs-菏麦13叶片1 h中刺激反应富集的基因数目明显较临麦2号叶片0 h-vs-菏麦13叶片0 h多,说明某些基因在高温条件下开始表达,对逆境做出响应。在分子功能方面,临麦2号叶片0 h-vs-临麦2号叶片1 h和菏麦13叶片0 h-vs-菏麦13叶片1 h结合物富集基因较多,临麦2号叶片0 h-vs-菏麦13叶片0 h和临麦2号叶片1 h-vs-菏麦13叶片1 h则是催化活性富集较多,表明高温胁迫下,基因以结合物的形式参与调控。临麦2号叶片1 h-vs-菏麦13叶片1 h在膜上富集的基因数目较临麦2号叶片0 h-vs-临麦2号叶片1 h少了许多,这可能是耐热品种与不耐热品种之间的差异。
3.2 不同品种在热胁迫下差异基因的KEGG分析
在KEGG 4种比较方案的分析中,临麦2号叶片0 h-vs-临麦2号叶片1 h和菏麦13叶片0 h-vs-菏麦13叶片1 h的差异基因显著富集的通路主要集中在内质网蛋白质加工、内吞、光合作用、谷胱甘肽代谢、剪接体等方面。光合作用发生次数最多,表明高温对光合作用的影响最大。光合速率的降低会阻碍同化物的运输,降低淀粉合成速率,导致小麦籽粒干瘪,千粒重减少,从而影响产量[1]。当高温胁迫发生时,植物的耐高温机制开始响应,如热激蛋白和热激转录因子开始参与高温胁迫,内质网蛋白质加工得到加强。内吞作用是钙离子进入细胞的途径之一,钙离子在抵抗高温胁迫方面也起着重要作用。有研究表明,在小麦叶片上喷洒一定浓度的CaCl2溶液可以有效提高小麦的耐热性,提高小麦产量[17]。谷胱甘肽参与诱导多种热激蛋白的表达,例如谷胱甘肽通过转录因子MYB21激活热休克蛋白基因bip3和hsp70b启动子,通过转录因子BZIP10激活hsp90.1启动子[18,19]。
3.3 部分转录因子对热胁迫的响应
转录因子是细胞响应外界胁迫以后主要调控某些基因转录的调控因子。朱丹等[20]以薄荷为试验材料,在光照培养箱设置40℃模拟高温胁迫处理2 h,采集叶片进行高通量测序,其结果显示与高温胁迫相关的转录因子家族主要集中在NAC、AP2-EREBP、WRKY、MYB、HSF等转录因子家族;张春霄[21]在黄花苜蓿中克隆到AP2/EREBP家族的MfERF049基因,对该基因进行了高温、低温、干旱、耐盐等非生物胁迫且该基因对上述非生物胁迫做出响应。李静宇等[22]证实MYB基因上启动子区域含有多种胁迫和生物激素的响应元件。NAC转录因子家族在调控包含高温在内的多种非生物胁迫时起到了非常重要的作用[23,24]。WRKY是植物中特有的一类转录因子家族,具有一个或两个WRKY结构域[25],与种子的休眠和萌发、作物的衰老和抵御逆境胁迫有关。HSF转录因子家族在对高温的响应中一直发挥着重要作用[26,27]。本研究结果发现在高温胁迫下,小麦旗叶中的转录因子包括NAC、AP2-EREBP、WRKY、MYB、HSF。这与前人结果一致,表明这些转录因子在小麦应对高温胁迫时发挥重要作用。该结果为后续探索小麦高温胁迫的分子机制及选育优质的耐高温品种提供理论依据。
