Selaginella pseudotamariscina (Selaginellaceae), an overlooked rosette-forming resurrection spikemoss from Vietnam
2022-11-16ZHANGMenghuaYANGJieLUUHongTruongTRANGioiZHANGXianchun
ZHANG Menghua, YANG Jie, LUU Hong Truong, TRAN Gioi, ZHANG Xianchun
( 1. State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China; 2. College of Life Sciences, University of Chinese Academy of Sciences, Beijing 100049, China; 3. Southern Institute of Ecology, Vietnam Academy of Science and Technology, Ho Chi Minh City 754000, Vietnam )
Abstract: A new species, Selaginella pseudotamariscina X. C. Zhang & C. W. Chen (Selaginellaceae), is described from Vietnam. The placement of this species within Selaginella subg. Stachygynandrum is assessed based on molecular phylogenetic analyses and morphological comparisons with related species. Molecular phylogenetic analyses suggest that S. pseudotamariscina is sister to S. digitata-S. imbricata clade. Morphologically, the new species is similar to S. tamariscina and S. pulvinata by sharing the rosette-forming habit, but distinguished by its dorsal leaves symmetrical, lanceate, sulcate on upper surface; strobili slightly dorsiventrally complanate and non-resupinate, sporophylls resembling vegetative leaves in form and arrangement, non-resupinate, the ventral sporophylls larger than the dorsal ones, dorsal sporophylls sterile, sporangia only borne on the base of ventral sporophylls.
Key words: chloroplast gene rbcL, resurrection plants, Selaginella subg. Stachygynandrum, new species, taxonomy
1 Introduction
SelaginellaP. Beauv. (1804) is the single genus of Selaginellaceae. It is the largest genus of lycophytes, containing ca. 700-800 species, widely distributed throughout the world, with the greatest diversity in the tropics and subtropics (Jermy, 1990; Weststrand & Korall, 2016a,b; Zhang et al., 2020). Several morphology-based classifications were proposed (Spring, 1840, 1850; Baker, 1883; Hieronymus & Sadebeck, 1902; Walton & Alston, 1938; Jermy, 1986). Based on the most recent molecular phylogenetic studies, seven subgenera were recognized within theSelaginella: subg.EricetorumJermy, subg.ExaltataeWeststrand & Korall, subg.Gymnogynum(P. Beauv.) Weststrand & Korall, subg.Lepidophyllae(Li Bing Zhang and X. M. Zhou) Weststrand & Korall, subg.RupestraeWeststrand & Korall, subg.Selaginella, and subg.Stachygynandrum(P. Beauv. ex Mirb.) Baker (Weststrand & Korall, 2016a, b; Zhang et al., 2020).
Selaginellais morphologically characterized by possessing rhizophores, heterospory, leaves generally arranged in four decussate rows, and terminal strobilus. Species of this genus have various growth forms, including climbing, creeping, prostrate, erect, suberect, and rosette forms (Jermy, 1990; Zhang, 2004; Zhang et al., 2013). Rosettes is a rare morphological feature inSelaginella(occurring in ca. 1% species). Only a few members of subg.Lepidophyllaeand subg.Stachygynandrumare rosettes which are all resurrection plants (Weststrand & Korall, 2016a).
Recently, Mr. Cheng-Wei Chen kindly shared us a new specimen (Wade5314) collected from the Southern central coast of Vietnam. It is different from S.tamariscina(P. Beauv.) Spring andS.pulvinata(Hook. & Grev.) Maxim. by its dorsiventrally complanate strobilus (vs. tetragonal strobilus) and symmetrical lanceate dorsal leaves. Historical collections from Vietnam of this species were all identified asSelaginellatamariscina(Alston, 1951). In order to correctly identify the specimen and infer the phylogenetic position of this unknown species, we analyzed its sequence of the chloroplast generbcLtogether with representatives of different subgenera ofSelaginellawith special attention to the rosettes plants. Both morphological and phylogenetic studies suggested that it represents an undescribed species. We therefore described it as a new species namedSelaginellapseudotamariscinaX.C. Zhang & C. W. Chen and presented the results here.
2 Materials and Methods
2.1 Morphological assessment
Morphologies of the newly collected specimens were examined and compared to those ofSelaginellatamariscinaandS.pulvinatausing specimens from the herbarium of Institute of Botany, CAS (PE). Sterile leaves, strobili, and sporophylls were observed and photographed under a Leica S9D stereo microscope.
2.2 Molecular analyses
In total, 32 individuals were sampled to represent 7 subgenera ofSelaginella, withIsoetesflaccidaas the outgroup. Sequences were obtained from the GenBank except for the new species (Voucher information and GenBank accession numbers are provided in the Table 1). Phylogenetic reconstruction was based on the chloroplast generbcL. We extracted total genomic DNA from silica gel dried materials using a modified cetyl trimethylammonium bromide (CTAB) method (Li et al., 2013). Library construction was performed with the NEBNext DNA Library Prep Kit (New England Biolabs, Ipswich, Massachusetts, USA). Paired-end reads of 2 × 150 bp were generated on an Illumina HiSeq 2500 instrument, andrbcLgene was extracted.
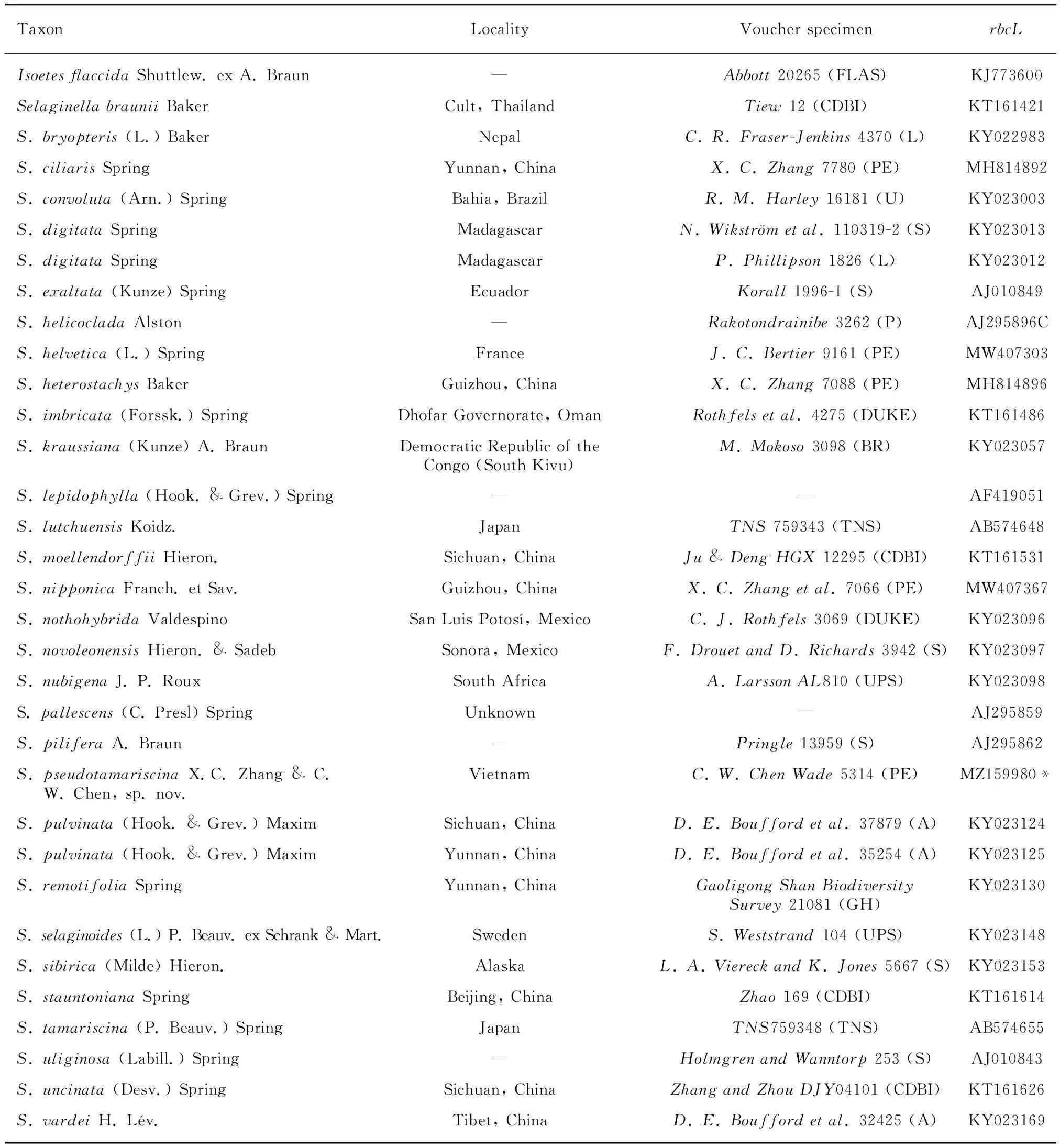
Table 1 Species names, localities, vocher information and GenBank accession numbers of DNA sequences used in this study
Sequences were aligned using MUSCLE (Edgar, 2004), followed by manual adjustment in PhyDE v0.9971 (Muller et al., 2010). Maximum likelihood (ML) analyses were performed using RAxML 7.2.6 (Stamatakis, 2006), with 1 000 bootstrap replicates under the GTRGAMMA model. We performed Bayesian inference (BI) analyses in MrBayes v. 3.2.6. (Ronquist et al., 2012) under the GTR + G + I model. For each Bayesian analysis, four MCMC chains were run simultaneously for 2 million generations, and sampled every 1 000 generations. The average standard deviation of split frequencies (<0.01) was used to assess the convergence. ML and BI trees and the branch support values were visualized using FigTree v.1.4.2 (Rambaut, 2014).

Table 2 Comparison of morphological characters of Selaginella pseudotamariscina, S. pulvinata, and S. tamariscina
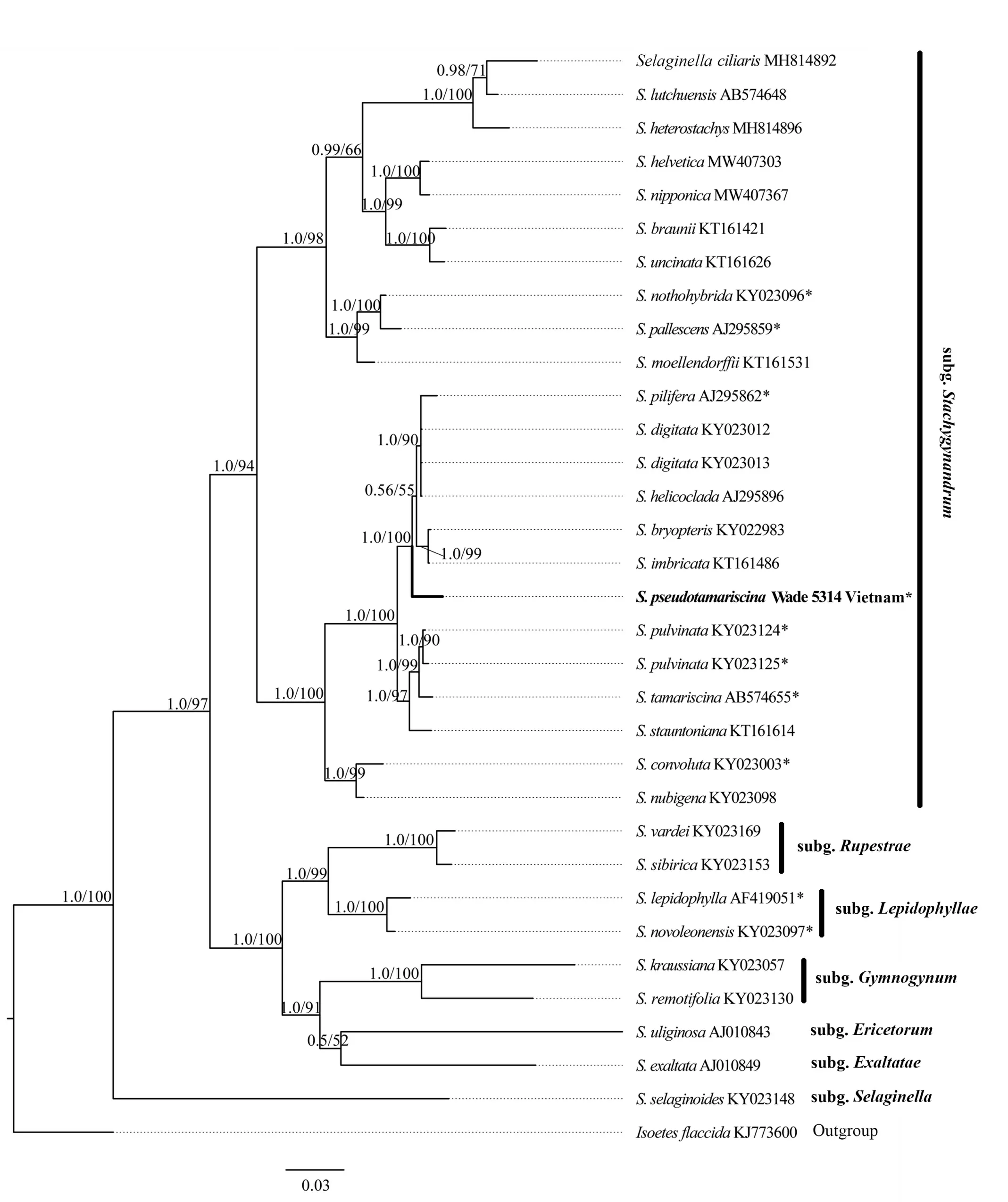
Bayesian inference posterior probability (PP) and maximum likelihood (ML) bootstrap are noted on the branches, respectively; The new species is shown in bold. * indicates rosette-forming.Fig. 1 Bayesian inference tree of the Selaginella pseudotamariscina and related species based on the rbcL gene

A. Individual; B. Upper view of branchlets; C. Lower view of branchlets; D. Axillary leaf (lower view); E. Ventral leaf (lower view); F. Dorsal leaf (upper view); G. Upper view of strobilus; H. Lower view of strobilus; I. Ventral sporophyll (lower view); J. Dorsal sporophyll (upper view).Fig. 2 Selaginella pseudotamariscina X. C. Zhang & C. W. Chen, sp. nov., Cheng-Wei Chen Wade 5314 (PE)

A1-C1. Selaginella pseudotamariscina (Vietnam, C. W. Chen Wade 5314, PE); A2-C2. S. pulvinata (China, Shanxi, Yellow River Exped. 251, PE); A3-C3. S. tamariscina (China, Fujian, X. C. Zhang et al. 9634, PE). A. Ventral leaves; B. Dorsal leaves; C. Axillary leaves.Fig. 3 Comparison of leaf morphology of Selaginella pseudotamariscina, S. pulvinata, and S. tamariscina
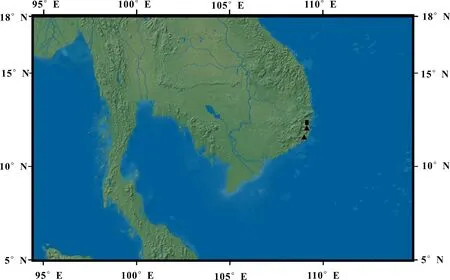
The rectangle represents the type location of S. pseudotamariscina.Fig. 4 Distribution of Selaginella pseudotamariscina X. C. Zhang & C. W. Chen, sp. nov.
3 Results and Discussion
The ML and BI topologies are totally identical, and the BI tree is shown in Fig. 1. The results of the molecular phylogenetic analyses showed that the new species nested within subg.Stachygynandrum, forming sister relationships with theS.pilifera—S.imbricataclade with weak support (PP=0.56/ML=55). However, of species nested in theS.pilifera—S.imbricataclade,S.piliferafrom America is the only rosette-forming species. TheS.pilifera—S.pseudotamariscinaclade is resolved to be sister to theS.tamariscina—S.stauntonianaclade with strong support (PP=1.0/ML=100). Morphologically, the new species is similar toS.tamariscinaandS.pulvinata. However, the new species differs obviously in several features, such as the slightly anisosporophylls which are similar to sterile leaves in form and arrangement; dorsal leaves symmetrical, lanceate, and sulcate on upper surface; ventral leaves shallowly sulcate on upper surface with their basiscopic margins ciliolate or denticulate; sporangia borne only on ventral side of strobilus axes ( Table 2; Fig. 2; Fig. 3: A1, B1).
4 Taxonomic Treatment
SelaginellapseudotamariscinaX.C. Zhang & C. W. Chen, sp. nov. (Fig. 2).
Type: VIETNAM. Khanh Hoa Province, Orchid Island, on granite rocks, in coastal open forest, 22 September 2018,Cheng-WeiChenWade5314 (holotype, SGN!; isotypes, PE!, SING!, TAIF!).
Diagnosis: The new species is similar toS.tamariscinaandS.pulvinatain the rosette-forming habits, differs by its slightly anisosporophyllous leaves which are similar to sterile leaves in form and arrangement, dorsal leaves symmetrical, lanceate, and sulcate on upper surface, ventral leaves shallowly sulcate on upper surface with their basiscopic margins ciliolate or denticulate, and only the ventral sporophylls fertile.
Description: Rosettes, xerophytic. Rhizophores restricted to basal portions of stems forming thick massive rootstocks; stems and roots entangled forming treelike trunk. Primary leafy branchlets 2-3 pinnately branched, branchlets compact and regularly arranged. Leaves thick, surfaces smooth. Axillary leaves on branches symmetrical, lanceate, or ovate-lanceate, ca. 2.18 × 0.73 mm, with membranous margins which measure ca. 1/2 of the width from margin to leaf midvein, lacerate or subentire, bases obtuse, and ciliolate, apice short aristate (ca. 0.02 mm long). Dorsal leaves strongly ascending, symmetrical, lanceate, 2.0-2.5 × 0.6-0.7 mm, sulcate on upper surface, carinate, bases obtuse, margins ciliolate or denticulate, slightly membranous, apices aristate (ca. 0.02 mm long). Ventral leaves slightly spreading, asymmetrical, ovate-lanceate to ovate-triangular, ca. 2.18 × 0.82 mm, shallowly sulcate on upper surface, apices aristate (ca. 0.04 mm long), basiscopic margins ciliolate or denticulate, acroscopic bases enlarged, broader than basiscopic, margins and becoming membranous outside, lacerate or subentire, ciliolate or lacerate proximally. Strobili solitary, terminal, compact, slightly dorsiventrally complanate, ca. 6 mm long; sporophylls similar to sterile leaves in form and arrangement, slightly anisophyllous; dorsal sporophylls smaller than ventral ones, lanceate-triangular, ca. 1.95 × 0.75 mm, margins ciliolate, slightly membranous, apices aristate (ca. 0.02 mm); ventral sporophylls triangular, ca. 2.13 × 0.98 mm, margins denticulate, ciliolate or lacerate, membranous, apices aristate (ca. 0.04 mm); only the ventral sporophylls fertile.
Additionalspecimensexamined: VIETNAM. Khanh Hoa Province, Nhatrang, Cau da, 100 m, 27 February 1922,Poilane2651 (P01244600, image online!; US01393274, image online!; VNM00021481!, VNM00021483!, VNM00021486!); Khanh Hoa Province, Nhatrang, 50 m, 10 September 1922,Poilane4529 (VNM00021477!, VNM00021479!, VNM00021482!); Khanh Hoa Province, Nhatrang, Nui Hon Heo, 3 May 1923,Poilane6173 (VNM00021478!); Ninh Thuan Province, Phan Rang, 27 February 1924,Poilane9768 (MICH1173518, image online!; VNM00021475!); Ninh Thuan Province, Phan Rang, 350 m, 3 March 1923,Poilane5541 (MICH1173519, image online!; VNM00021480!); Ninh Thuan Province, Phan Rang, 200 m, 6 March 1923,Poilane5616 (P01244598, image online!; VNM00021484!).
Distributionandhabitat:Selaginellapseudota-mariscinais only known in Khanh Hoa and Ninh Thuan provinces of southern central Vietnam (Fig. 4), growing on granite rocks in open coastal forests.
Etymology: The specific epithet ‘pseudotamari-scina’ refers to its close relation and morphological similarity withS.tamariscina.
Conservationstatus(EN):Selaginellapseudotamariscinais distributed only in two provinces in southern central Vietnam (Khanh Hoa and Ninh Thuan).
There exist a few specimens ofS.pseudotamariscinadeposited in various herbaria and one of the oldest specimens was collected a century ago by the French collector E. Poilane in Nhatrang, which is preserved in herbarium P with a duplicate in the U.S. This rosette-forming species appears to have adapted to the coastal climate on granite rocks in Southern central Vietnam. Here it is tentatively listed as an endangered (EN) species according to IUCN categories and criteria (IUCN, 2018).
KeytoSelaginellapseudotamariscina,S.tamariscinaandS.pulvinata
1. Strobilus dorsoventrally complanate; ventral sporophylls fertile, dorsal sporophylls sterile; dorsal leaves symmetrical, lanceate, sulcate on the upper surface; ventral leaves shallowly sulcate on the upper surface
S.pseudotamariscinaX. C. Zhang & C. W. Chen
1. Strobilus tetragonal; ventral and dorsal sporophylls both fertile; dorsal leaves asymmetrical, obliquely ovate, triangular, or elliptic, upper surface without sulcate; ventral leaves upper surface without sulcate
2
2. Stems and roots entangled not forming treelike trunk; inner margins of dorsal leaves lacerate or entire, outer margin revolute and entire
S.pulvinata(Hook. & Grev.) Maxim.
2. Stems and roots entangled often forming treelike trunk; inner margins of dorsal leaves denticulate, outer margin denticulate or shortly ciliolate
S.tamariscina(P. Beauv.) Spring
AcknowledgementsWe thank Van Son DANG, the curator of VNM for providing the access to their collections and Taiwan Pteridophyte Research Group (www.twfern.org) for sharing the silica-dried material.
