福建山美水库浮游动物群落结构时空特征及其影响因子分析*
2022-11-09刘智暘王江滨廖传松李俊锋王靖斌熊满堂张堂林郭传波刘家寿
刘 俏,刘智暘,王江滨, 廖传松,李俊锋,4,郭 超,4,王靖斌,熊满堂,4,张堂林,4,郭传波,4,刘家寿,4**
(1:华中农业大学水产学院,武汉 430070) (2:中国科学院水生生物研究所,淡水生态与生物技术国家重点实验室,东湖湖泊生态系统试验站,武汉 430072) (3:福建省泉州市山美水库水资源调配中心,泉州 362008) (4:中国科学院大学,北京 100049)
浮游动物是水生态系统中的重要初级消费者,以浮游植物、碎屑、细菌和腐殖质等为食,在食物网中对控制浮游植物密度和群落结构具有重要作用[1-2],也具有净化水质的功能[3]. 同时,浮游动物的生命周期为几天到几个月不等,发育快,代谢旺盛,是水体中滤食性鱼类、大多数小型鱼类和处于生活史早期阶段鱼类的天然优质饵料[4]. 浮游动物在联系初级生产者和高营养级消费者方面起到承上启下的作用,在水生生态系统物质循环、能量流动过程中也具有至关重要的作用[5]. 此外,浮游动物群落结构也受到水体生物和非生物因素的影响,对水环境变化的响应非常敏感,可以作为水质状况的指示种. 例如,螺形龟甲轮虫(Keratellacochlearis)、萼花臂尾轮虫(Brachionuscalyciflorus)、角突臂尾轮虫(Brachionusangularis)等为富营养类水体指示种,广布多肢轮虫(Polyarthravulgaris)等为中营养类指示种,异尾轮属(Trichocerca)等为贫营养类指示种[6-7]. 改善浮游动物群落结构、提升浮游动物密度和生物量对于维护水生态系统健康状况与水质保护具有重要意义.
鱼类是水生态系统中的顶级消费者,参与水域食物网物质循环和能量流动,对于水生态系统结构与功能调节具有重要作用[8]. 鱼类具有多种摄食习性,不同食性类型的鱼类对浮游动物群落具有不同的影响,是水生态系统中影响浮游动物群落的重要生物因素. 一方面,典型滤食性鱼类,如鲢(Hypophthalmichthysmolitrix)和鳙(Aristichthysnobilis)的滤食行为可一定程度影响浮游动物的生物量[9];另一方面,湖泊、水库中的小型鱼类如(Hemiculterleucisculus)、似鱎(Toxabramisswinhonis)、飘鱼(Pseudolaubucasinensis)等大多以大型浮游甲壳动物为主要食物,通过加强鱼食性鱼类对这些小型鱼类的捕食,改变其对大型浮游甲壳动物的摄食强度,进而增加水体中大型浮游甲壳动物的生物量,也是经典生物操纵的基本原理[10]. 湖泊、水库中浮游动物群落与鱼类及水环境的关系一直是水域生态学研究的热点[11].
山美水库(25°07′41″N,118°26′36″E)位于福建省泉州市西北部的南安市,因大坝位于九都镇山美村而得名,集水面积23.75 km2,总库容6.55亿m3,正常蓄水位高96.48 m,全年平均水深15.3~46.6 m[12],是晋江流域内唯一一座集防洪、供水、灌溉、发电等综合利用于一体的大型水利枢纽工程[13]. 山美水库不仅是泉州市重要水源地,保障着400多万人口的生产生活用水,灌溉面积4.07万hm2,还肩负着为台湾省金门县供水的任务,因此维护山美水库水生态系统健康至关重要[14-15]. 2010年对于山美水库水质的研究表明,总氮浓度(2.23~4.91 mg/L)、总磷浓度(0.007~0.132 mg/L)较高[16],2012年春季和夏初在入库区底泥淤积严重区域进行了清淤疏浚措施,有效改善了水质[17],但2017年山美水库月平均总氮浓度为2.15 mg/L[18],且近年来山美水库春、夏季pH值常偏高. 另外,关于山美水库水质与重金属含量[19],氮、磷含量[20-21],沉积物中有机物含量[22-23]等已有一些研究,这些研究表明山美水库水质呈现高氮低磷(130∶1)的特征,沉积物中的氮、磷及有机物主要源于入库支流. 相对而言,对于山美水库水生生物方面的关注较少,关于浮游动物群落结构及其与环境因子的关系缺乏研究. 为了提高山美水库浮游动物特别是大型浮游甲壳动物的生物量,减少山美水库浮游植物的生物量,减轻光合作用强度进而降低pH值,2020年以来,结合经典生物操纵与非经典生物操纵方法,调整了山美水库鱼类放养策略和捕捞措施,一是放养以鲢为主的滤食性鱼类以控制藻类密度,二是放养以翘嘴鲌(Culteralburnus)、花鲈(Lateolabraxjaponicus)和南方鲇(Silurusasotus)为主的鱼食性鱼类以控制小型鱼类进而壮大浮游动物类群,以及以黄尾鲴(Xenocyprisdavidi)为主的碎屑食性鱼类以减少水体中悬浮有机颗粒含量. 本研究通过2020-2021连续两年逐季调查山美水库鱼类调控后浮游动物群落结构及水环境状况,分析浮游动物群落结构时空动态及其与水环境因子的关系,以期为湖泊、水库的水生态系统管理和调控提供科学依据.

图1 山美水库浮游动物与水环境采样点分布Fig.1 The distribution of zooplankton and water environment sampling sitesin the Shanmei Reservoir
1 材料与方法
1.1 采样点分布及样品采集
在山美水库共设置21个采样点,根据采样点在库区的空间分布,将其分成4个水域:1~5号点和21号点为入库(A1),18、19、20号点为库首(A2),6、7、8号与12~17号点为库中(A3),9、10、11号点为库尾(A4) (图1). 分别于2020年和2021年的1、4、7和10月采集浮游动物样品和水样.
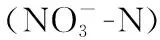
浮游动物定性样品采用25号浮游生物网(孔径64 μm)在表层水体做“∞”状采集3~5 min,将样品装入50 mL聚乙烯瓶,现场加入4%甲醛溶液固定,后带回实验室参照文献[25-29]进行鉴定. 原生动物和轮虫定量样品用10 L采水器分层采集30 L水样,充分混合,取其混合水样1 L,加入1%鲁哥氏液固定,然后带回实验室经沉淀48 h后浓缩至100 mL左右,再次沉淀48 h后浓缩至50 mL,加入4%甲醛保存. 浮游甲壳动物用5 L采水器分层采集30 L水样,用13号浮游生物网(孔径112 μm)过滤至50 mL样品瓶中,并加4%甲醛溶液将其固定. 在显微镜下对所有的浮游动物进行鉴定、计数,密度和生物量的计算参照文献[25-30].
2020年起初,山美水库管理部门开展了基于鱼类放养和捕捞调控的鱼类群落调控,主要对象包括滤食性、鱼食性和碎屑食性等鱼类,于2020年1-3月、2021年1月和2月放养鱼类,除2020年12月外山美水库逐月开展鱼类捕捞,其中2021年4-6月未捕捞鲢鳙. 鱼类放养和捕捞数据见表1.
表1 2019-2021年山美水库鱼类放养和捕捞数据
Tab.1 Stocking and fishing data of fishes in the Shanmei Reservoir during 2019-2021

鱼类种类放养量/t捕捞量/t2019年2020年2021年2019年2020年2021年鲢Hypophthalmichthys molitrix13.60135.2136.572.8527.27111.68鳙Aristichthys nobilis63.2976.478.3218.05150.88128.83翘嘴鲌Culter alburnus01.500.490.350.60鳜Siniperca chuatsi00.250.067000南方鲇Silurus asotus01.41.3000花鲈Lateolabrax japonicus00.530.642.580.843.95罗非鱼Tilapia00018.4417.5984.04黄尾鲴Xenocypris davidi04.33.5000其它00.5201.160.511.22总76.89220.1220.307313.57197.44330.32
1.2 数据处理与分析
采用线性混合效应模型(linear mixed effects model, LMM)比较各水质指标在不同水域、季节间的差异,采用Emmeans函数进行两两比较. LMM用于检验浮游动物生物量和密度在不同季节、水域间的差异,以季节和水域作为固定效应因素,采样点和采样年份作为随机因素. 采用优势度指数(Y)来评估优势种,Y>0.02的物种为优势种[31]. 采用Shannon-Wiener 多样性指数(H′)、Pielou均匀度指数(J)以及Margalef丰富度指数(D)[32]分析浮游动物群落的多样性特征,计算公式分别如下[33-34]:
(1)
(2)
(3)
(4)
式中,ni为第i个物种的密度,N为全部物种的总密度,fi为第i个物种的出现频率,S为总种类数.
以多样性指数评估水质的标准如下[35]:H′:0~1为多污型,1~2为α-中污型,2~3为β-中污型,>3为清洁-寡污型;D:0~1为多污型,1~2为α-中污型,2~3为 β-中污型,3~4为寡污型,>4为清洁型;J:0~0.3为多污型,0.3~0.5 为中污型,0.5~0.8为寡污型,0.8~1.0为清洁型.
浮游动物密度和生物量可作为表征湖泊营养程度的生物指标之一[36-39],判断依据为密度<1000 ind./L为贫营养,1000~3000 ind./L为中营养,>3000 ind./L为富营养;生物量<1 mg/L为贫营养,1~3 mg/L为中营养,>3 mg/L为富营养.
筛选出至少在一个采样点的密度占比≥1%,且出现频率≥30%的浮游动物种类进行去趋势对应分析(detrended correspondence analysis,DCA)[40-41]. 共筛选出35种浮游动物进行分析,由于DCA分析中最大梯度长度大于4,因而选择典范对应分析(canonical correspondence analysis, CCA)探究浮游动物与环境因子之间的关系[42],经共线性分析后,选用电导率、水温、硝态氮、溶解氧、透明度、pH、水深、总氮、叶绿素a和高锰酸盐指数水环境因子分析其与浮游动物群落结构的关系,浮游动物种类的代码见附表Ⅰ.
采用R软件“lme4”和“Emmeans”包进行线性混合效应模型分析与两两比较,Canoco 5.0软件进行去趋势对应分析与典范对应分析,P<0.05视为差异显著,P<0.01视为差异极显著,采用ArcGIS 10.6软件制作采样点图.
2 结果与分析
2.1 水体理化特征
山美水库水体理化因子在不同季节、不同水域的变化如表2和表3. 从季节上来看,山美水库所有水质指标均呈现显著的季节变化(LMM,P<0.05;表2,图2). 夏、秋季总磷浓度显著高于春、冬季;夏季总氮浓度显著高于秋季;夏季氨氮浓度显著低于秋季;春季和冬季硝态氮浓度显著高于夏秋季;高锰酸盐指数、叶绿素a和溶解氧浓度在春夏季显著高于秋冬季;秋冬季透明度显著高于春夏季;夏季pH值最高,秋季最低.

表2 山美水库水体理化指标季节变化*
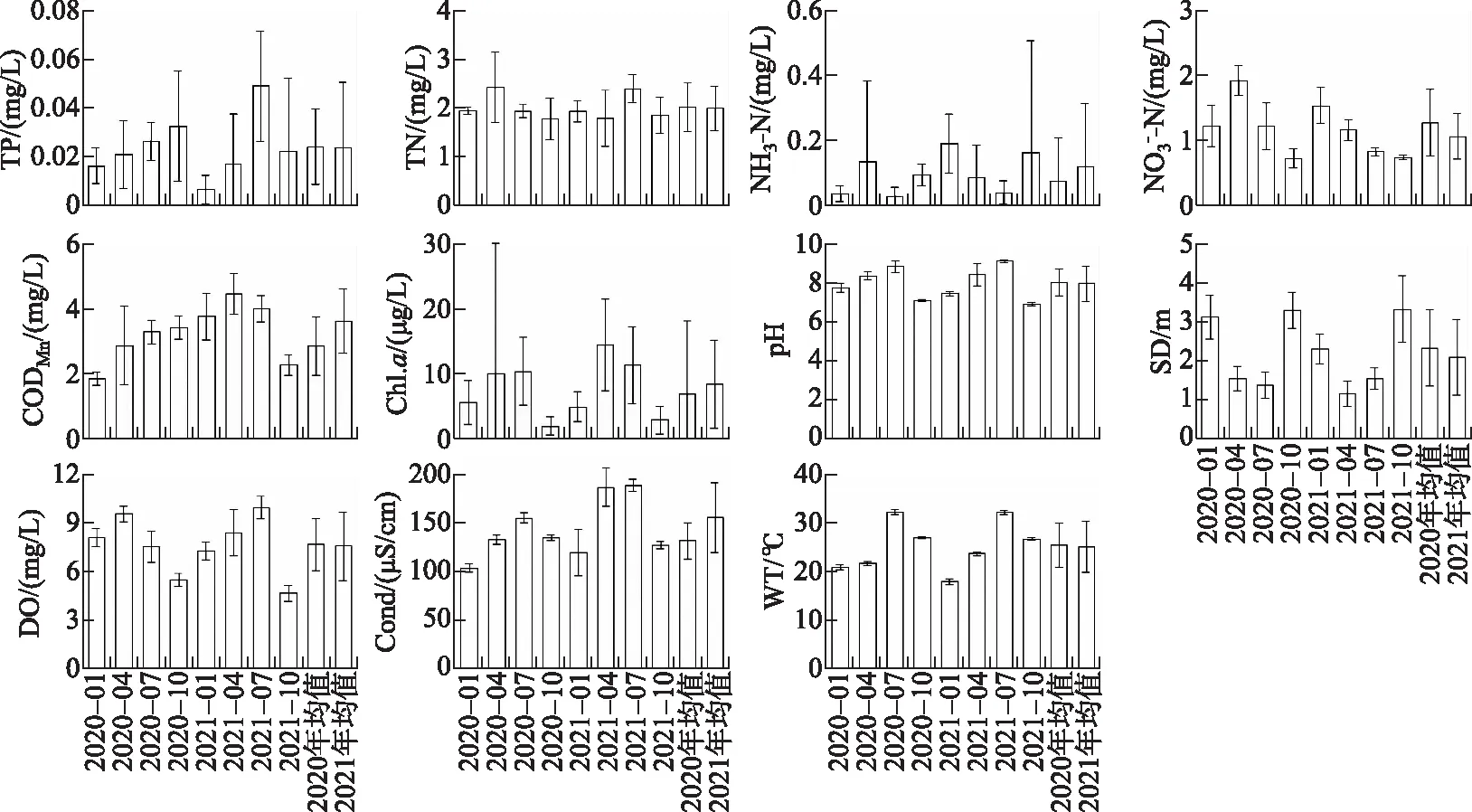
图2 山美水库水体理化指标季节变化Fig.2 Seasonal changes of main physical and chemical indicators of the Shanmei Reservoir
从不同水域间水体理化指标比较来看,总磷、氨氮、电导率、硝态氮和水温在4个水域间无显著性差异(LMM,P≥0.05;表3). 高锰酸盐指数、叶绿素a在入库水域的浓度显著高于库中,pH值、透明度和溶解氧在入库水域的值显著低于其他区域,其他区域间无显著性差异.
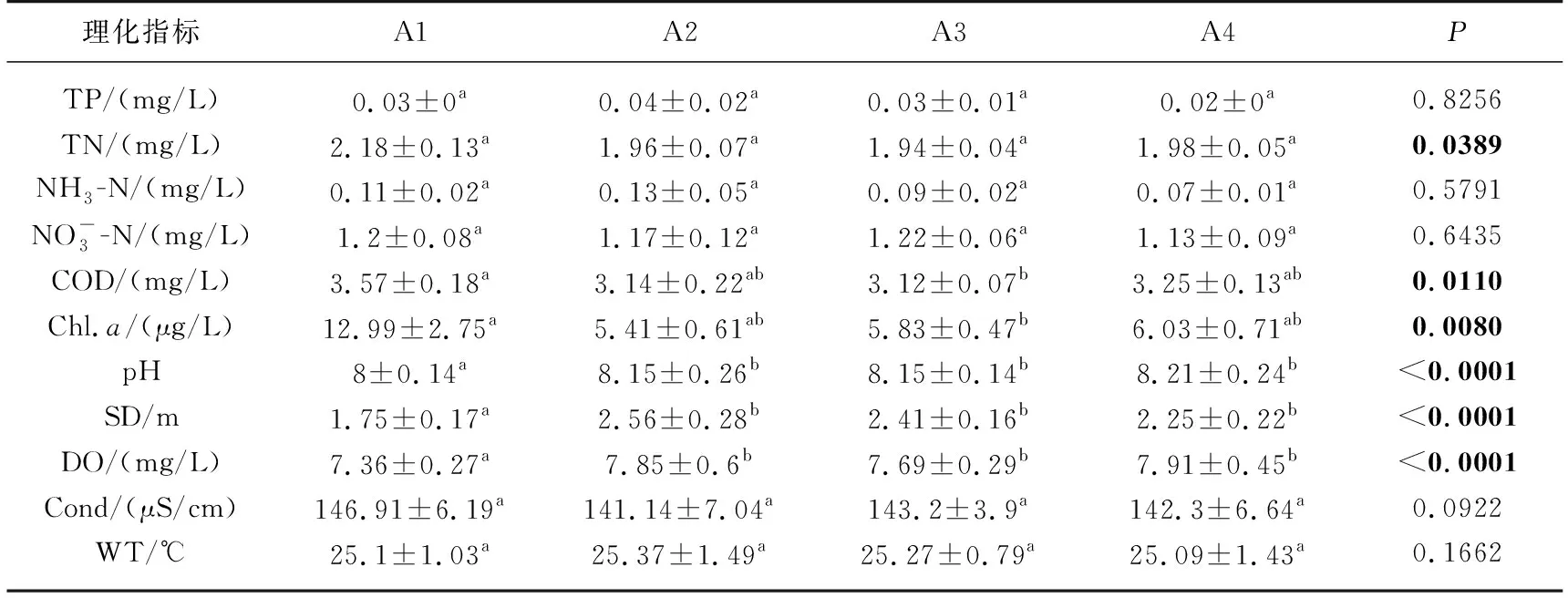
表3 山美水库水体理化指标空间变化*
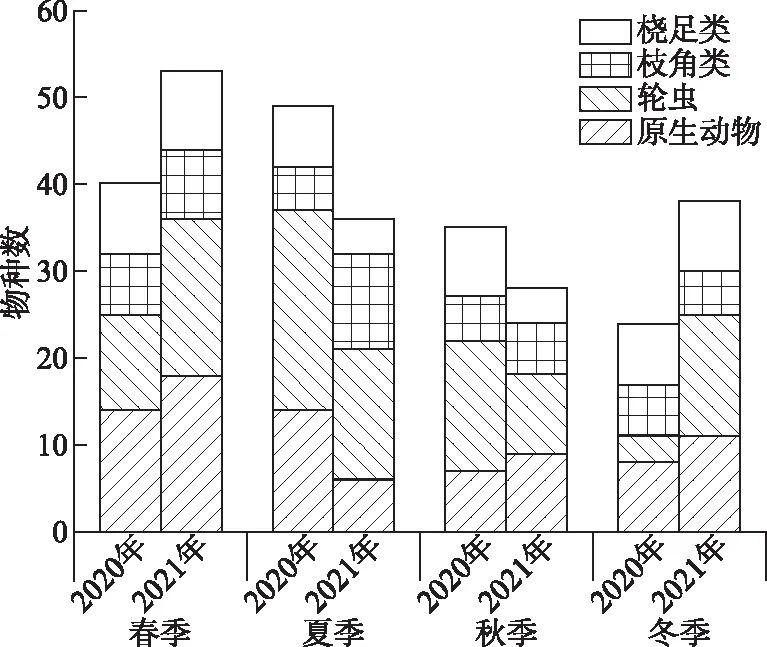
图3 山美水库浮游动物4个类群种类数季节变化Fig.3 Seasonal variation in percentages of four categories of zooplankton in the Shanmei Reservoir

表4 山美水库浮游动物密度时空比较*
2.2 浮游动物群落结构
2.2.1 物种组成 2020-2021年共鉴定浮游动物4门68属102种,其中轮虫21属41种,占总种类数的40.20%;原生动物32属41种,占40.20%;枝角类8属13种,占12.75%;桡足类7属7种,占6.86% (附表Ⅱ;图3). 2021年春、冬季种类数(分别为38和53种)均多于2020年春、冬季(分别为24和40种)种类数,2020年夏、秋季种类数(分别为49和35种)均多于2021年夏、秋季(分别为36和28种).
2.2.2 密度和生物量 2020-2021年山美水库浮游动物平均密度为(1443.05±360.02) ind./L,变化范围为418.72~3592.74 ind./L. 其中,原生动物的平均密度为(789.38±324.65) ind./L,轮虫为(549.6±199.82) ind./L,枝角类和桡足类分别为(18.19±6.26)和(85.89±24.82) ind./L.
山美水库浮游动物密度在不同季节差异显著(P<0.05),春季浮游动物密度显著高于其他季节,其它季节间无显著差异. 其中,枝角类在夏季的密度显著高于其它季节,春季显著高于冬季;桡足类在秋季的密度显著高于其它季节;原生动物在春季的密度显著高于夏秋季,冬季显著高于夏季;轮虫在夏季的密度显著高于其它季节,春季显著高于冬季. 从年度上来看,2021年浮游动物密度((2007.97±574.97) ind./L)显著高于2020年((878.14±248.04) ind./L),2021年四大类浮游动物的密度分别显著高于2020年四大类的密度(表4,图4).
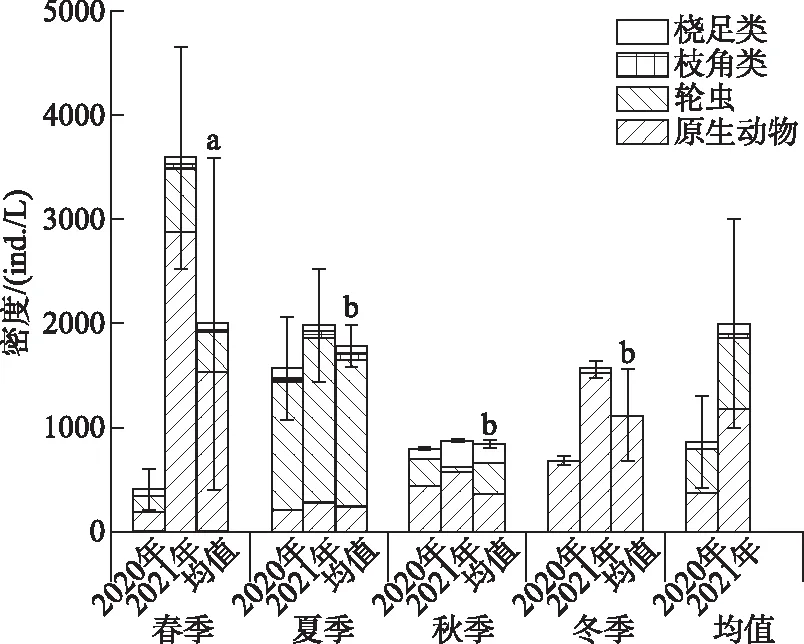
图4 山美水库浮游动物平均密度季节及年度变化(字母不同表示差异显著)Fig.4 Seasonal and annual variations of zooplankton density in the Shanmei Reservoir(Difference in lowercase indicates significant difference in seasons)
2020-2021年山美水库浮游动物平均生物量为(1.21±0.27) mg/L,变化范围为0.19~2.34 mg/L. 其中,原生动物的平均生物量为(0.04±0.02) mg/L,轮虫为(0.52±0.17) mg/L,枝角类和桡足类分别为(0.29±0.07)和(0.36±0.12) mg/L.
山美水库浮游动物生物量在季节间差异极显著(P<0.01),夏季显著高于其他季节,春秋季显著高于冬季,春季和秋季之间差异不显著(P>0.05). 其中,枝角类在夏季的生物量显著高于其它季节,春秋季显著高于冬季;桡足类在秋季的生物量显著高于其他季节;原生动物在春季的生物量显著高于夏秋季,冬季显著高于夏季;轮虫在夏季的生物量显著高于其他季节,春季显著高于冬季. 从年度上来看,2021年浮游动物生物量((1.43±0.28) mg/L)显著高于2020年((0.99±0.48) mg/L),其中,枝角类生物量2021年A2、A3和A4区域显著高于2020年对应区域;桡足类和原生动物生物量2021年显著高于2020年;轮虫生物量年际间无显著差异(表5,图6).
表5 山美水库浮游动物生物量时空比较*

浮游动物生物量在区域间差异显著(P<0.05),入库区域((1.44±0.31) mg/L)显著高于库中((1.06±0.22) mg/L)和库尾区域((1.09±0.19) mg/L),库中、库尾和库首区域之间的生物量差异不显著(P>0.05)(图7).
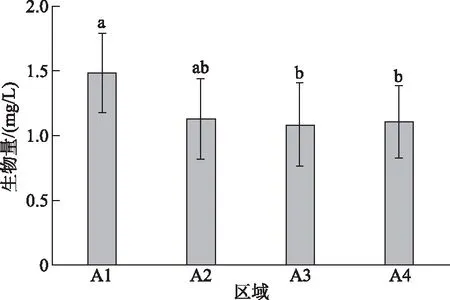
图7 山美水库浮游动物生物量区域比较(字母不同表示差异显著)Fig.7 Spatial comparison of zooplankton biomass in the Shanmei Reservoir(Difference in lowercase indicates significant difference in areas)
2.2.3 多样性特征 浮游动物优势种存在明显季节差异,夏季优势种组成最复杂,其他季节相对简单. 原生动物优势种主要出现在冬季、春季和秋季,其中王氏似铃壳虫(Tintinnopsiswangi)在这3个季节均出现过,其它原生动物仅出现在单个季节或2个季节;轮虫优势种主要出现在夏季,其中暗小异尾轮虫(Trichocercapusilla)、卵形无柄轮虫(Ascomorphaovalis)以及疣毛轮属(Synchaetasp.)在春秋两季也有出现,广布多肢轮虫出现在夏季、秋季和冬季;哲水蚤桡足幼体为2021年秋季优势种,剑水蚤桡足幼体为多季度优势种(表6). 各个季节的优势种所属类群有所变化,春、冬季的优势种主要为原生动物,夏、秋季主要为轮虫.

表6 山美水库浮游动物优势种季度变化
山美水库浮游动物多样性指数如图8,Margalef丰富度指数范围为1.97~3.01,平均值为2.46,对应水质为β-中污染;Pielou 均匀度指数范围为0.42~0.75,平均值为0.62,对应水质为轻污染;Shannon-Wiener 多样性指数范围为1.10~2.31,平均值为1.75,对应水质为α-中污染.
对3个多样性指数进行混合效应模型分析发现,2021年3个指数的平均值均显著高于2020年. 山美水库的水质状况为轻~中度污染.

图8 山美水库浮游动物多样性指数区域和季节变化Fig.8 Spatial and seasonal variations of zooplankton diversity index in the Shanmei Reservoir
2.3 浮游动物群落结构与环境因子的关系
CCA分析的第一轴和全部轴均为差异极显著(Monte Carlo Test,P<0.01). CCA结果显示,环境变量对响应变量总解释率为59.1%,调整后的R2为52.2%;第一轴和第二轴特征值分别为0.8328和0.4147,共解释了浮游动物群落变异程度的39.08%. 经蒙特卡洛置换检验,电导率、水温、硝态氮、溶解氧、透明度和高锰酸盐指数对浮游动物群落结构影响极显著(P<0.01),pH、水深、总氮和叶绿素a浓度对浮游动物群落结构有显著影响(P<0.05),氨氮和总磷的影响不显著(P>0.05).
不同季节间,影响筛选的浮游动物物种的主要环境因子有所差别. 春季砂壳虫属、卵形无柄轮虫、溞属、汤匙华哲水蚤、舌状叶镖水蚤、台湾温剑水蚤喜好出现,与溶解氧、硝态氮浓度呈正相关,与总氮浓度呈负相关;秋季僧帽溞、跨立小剑水蚤、大型中镖水蚤与透明度、总氮浓度呈正相关;冬季似铃壳虫、微型裸腹溞、剑水蚤桡足幼体和右突新镖水蚤与透明度和溶解氧浓度呈正相关,与其他影响因子呈负相关,但透明度是主要影响因子;夏季在水温、电导率、pH、高锰酸盐指数相对较高的环境下,其余浮游动物成为喜好出现的物种,这些物种与透明度、溶解氧浓度、硝态氮浓度呈负相关(图9).

图9 山美水库浮游动物与环境因子的CCA排序图(绿色:春季;黄色:夏季;红色:秋季;紫色:冬季;圆形:采样点;三角形:物种)Fig.9 CCA of the zooplankton and associated environmental factors in the Shanmei Reservoir(Green: Spring; Yellow: Summer; Red: Autumn; Purple: Winter; Circular: site; Triangle: species)
3 讨论
3.1 山美水库水质时空变化
山美水库水体理化因子呈现明显季节变化,夏季总磷、总氮、pH值、电导率、水温较高,春季高锰酸盐指数、叶绿素a和溶解氧浓度最高,冬季、秋季透明度较高. 夏季和秋季总磷浓度较高可能是由于浮游植物的光合作用加强,pH升高,促进了磷的释放[43-44],另外水温的升高也能促进夏季和秋季水库磷的内源释放[45],电导率主要反映水体中可溶性离子总量,研究表明,温度是影响电导率的主要因素,在一定温度范围内,电导率与水温呈显著正相关关系[46]. 叶绿素a和溶解氧浓度在春季较高,这可能是因为春季水温回升浮游植物大量繁殖,叶绿素a浓度升高,光合作用加强,溶解氧升高;同时,浮游植物春夏季光合作用的加强造成了pH值的升高,这是山美水库水质面临的主要问题之一,从本研究结果来看,2021年夏季pH值相较于2020年无明显下降,这可能是由于获得的数据是每个季度单次调查所得,无法全面反映山美水库pH值的变化情况. 然而,从山美水库管理中心国控点长期实时监测的数据来看,2021年pH值的峰值(9.48)较2020年(9.70)有所降低,且全年pH值超过9.0的天数由2020年的112天缩短至2021年的68天(内部资料),表明夏季pH偏高问题有所缓解,这可能是由于山美水库藻类得到了一定控制,从而降低了光合作用强度的缘故.
水质的空间变化方面,山美水库入库区域的溶解氧、pH值和透明度显著低于其他区域,高锰酸盐指数和叶绿素a浓度则显著高于其他区域,总体呈现出入库区域水质劣于其他区域的特征. 这种水质的空间格局在水库生态系统中很常见,例如官厅水库[47]、茜坑水库[48]和青草沙水库[49]等. 山美水库上游2条入库支流受生产生活的影响[21],营养盐浓度较高(总氮高42.34%,总磷高70.85%;未发表数据),在入库区域大量沉积,是导致该区域水质劣于其他水域的主要原因. 因此,尽管由于营养盐的沉积导致此区域浮游植物大量繁殖,但是此区域水体较浅且流速较快,尤其是在低水位季节,浮游植物的光合作用受到影响,CO2利用率低于库区,且浮游植物死亡后的有氧分解也会导致该区域的溶解氧消耗高于库区,从而产生大量的CO2,因此该区域的溶解氧和pH显著低于其他区域[50].
3.2 浮游动物群落结构的时空特征
山美水库的浮游动物种类组成以轮虫为主,其次是原生动物,枝角类和桡足类较少,这与陈昕等[35]、陈莎等[51]、严广寒等[52]、刘盼盼等[53]在其他水域的研究结果一致. 整个调查期间,山美水库浮游动物种类组成、优势种、密度和生物量存在季节差异. 水温是影响浮游动物生长的重要因子[54],夏季水温高有利于水中的物质循环和能量流动以及浮游植物等的大量繁殖[35],从而间接影响到浮游动物的捕食活动[55],促进浮游动物大量繁殖. 全年优势种中,轮虫和原生动物等小个体类群占比较高,说明山美水库的浮游动物群落呈现小型化特征,这可能与水库优势鱼类种群——似鱎、等对桡足类、枝角类的捕食有关[56]. 同时,温度也是影响浮游动物生长的重要因素,春季、冬季山美水库水温(20~22℃)正好处于原生动物最适生长温度区间[57],夏季、秋季水温(26~32℃)更适合轮虫生长,轮虫的休眠卵大量孵化,轮虫种群周转加快,导致种群密度迅速增加[30].
从浮游动物群落的多样性指数与浮游动物密度和生物量来看,山美水库水质整体处于轻度~中度污染状态,与之对应的是,山美水库夏季优势度最大的暗小异尾轮虫以及其他异尾轮属、胶鞘轮属的轮虫为寡污型指示种[39,56],锯齿龟纹轮虫、广布多肢轮虫等均为中污染型水体的常见种[39,56-58],春季、秋季和冬季的优势种多为寡污性的砂壳虫属和似铃壳虫属的种类[36,39,56,59]. 这些说明了夏季山美水库处于中营养状态,春秋冬季处于贫营养状态,另外,尽管2020和2021年山美水库浮游动物群落结构多样性3个指数所对应的水质类型一致,但2021年的3个指数平均值均高于2020年,这表明山美水库浮游动物多样性和丰富度有所提高,水质得到改善[60].
3.3 浮游动物群落结构与水环境因子的关系
水库中浮游生物、鱼类多样性高,水态系统结构复杂,浮游动物群落结构受非生物因素和种间相互作用的影响[2]. 研究表明水温是影响浮游动物季节变化的主要水环境因子,例如,对长江干流浮游动物群落结构与水环境因子分析表明,水温与轮虫、枝角类和桡足类的密度和生物量呈正相关关系[61]. 本研究中,大部分轮虫种类喜好水温、电导率、pH值和高锰酸盐指数较高的夏季水环境;枝角类和桡足类最适生长温度为20~30℃[62],在春秋季优势度较高;原生动物则喜好出现在冬季. 同时,由于似铃壳虫属多为清洁型种类,冬季的透明度是影响浮游动物群落结构的主要因子.
除水温外,电导率、溶解氧、氮、磷以及高锰酸盐指数等也是影响浮游动物分布的重要水环境因子[63-64],浮游动物虽然不直接依赖营养物质生存,但受水库中浮游植物、细菌和碎屑的影响,水库营养状态仍然可能影响浮游动物丰度、群落结构、体型和生产力[65-66]. 本研究中,电导率、硝态氮、溶解氧和高锰酸盐指数是影响浮游动物分布的主要营养盐指标. 春季影响浮游动物分布的指标主要是硝态氮和溶解氧,这可能是由于硝态氮在一定程度上为浮游植物的生长提供了氮源[67],从而间接为浮游动物提供了食物来源. 溶解氧是浮游动物生存的重要因子之一,研究显示过高的溶解氧会抑制一些轮虫和枝角类的生长[68]. 在本研究中,春季山美水库平均溶解氧浓度最高,浮游动物密度与溶解氧浓度呈正相关,主要原因可能是因为这些浮游动物的生长需要较高的溶解氧,而在其他季节溶解氧不是浮游动物密度的限制因子. 夏季浮游动物密度和生物量与高锰酸盐指数、电导率和总磷等多种营养盐呈正相关,说明夏季浮游动物能在较高营养水平水域生存,耐污能力较强.
3.4 鱼类群落调整与改善浮游动物群落及水质的关系
枝角类和桡足类在水生食物网中对于浮游植物、细菌和悬浮物具有很强的摄食能力,对于浮游植物群落结构的改变起到重要影响,从而可以达到降低水库浮游植物生物量等目的[69-70]. 因此,调控山美水库浮游动物的群落结构、提高大型浮游甲壳动物的比例对改善水库水质具有重要作用.
本研究采用组合调控措施来优化山美水库浮游动物群落结构. 组合调控措施实施后,浮游动物群落结构发生显著变化. 2021年山美水库浮游动物密度和生物量均显著高于2020年,其中,枝角类、桡足类的密度和生物量也显著提升,考虑到2021年鱼类捕捞量是2020年的1.67倍,推测鳙等浮游动物食性鱼类的生物量可能是影响浮游动物密度和生物量的重要原因之一[71-72]. 同时本研究也发现,2021年桡足类优势种较2020年的剑水蚤幼体增加了哲水蚤幼体,而剑水蚤无节幼体和哲水蚤主要以藻类为食[73],说明山美水库桡足类的种群得到了一定的恢复,而桡足类为了躲避鱼类的捕食会减少成年个体,转而以活动能力更强的幼体为主要形式在水体中出现[74];优势种中未出现枝角类,可能是由于在鲢鳙的摄食压力之下,枝角类对于鲢鳙的摄食更加敏感,大型枝角类的种群会受到一定影响[75]. 另一方面,在湖泊、水库中放养鱼食性鱼类也能通过食物网级联效应影响浮游动物群落结构,这是经典生物操纵理论的基础[77],也在中国得到证实. 例如,Peng等[76]在泉州龙湖放养不同食性鱼类发现,放养鱼食性鱼类实验组(花鲈等)的浮游动物生物量高于放养滤食性鱼类和放养杂食性鱼类实验组. 目前山美水库小型鱼类(似鱎、等)较多(相对丰度76.05%,相对生物量37.74%;未发表数据),这些小型鱼类多以大型浮游动物为饵料[77]. 鉴于此,相较于2020年前,2020年和2021年在山美水库增加了翘嘴鲌、花鲈和南方鲇等鱼食性鱼类的放养,加强对这些小型鱼类的捕食强度,也是大型浮游甲壳动物密度和生物量得以提高的因素之一. 因此,从水质管理与调控的角度,应该加大对小型鱼类和浮游动物食性鱼类的捕捞强度,同时适当放养鱼食性鱼类通过下行效应控制小型鱼类密度[78],既可改善水质,也能减轻小型鱼类对经济鱼类的食物竞争、提高饵料资源的利用[79]. 由于经典生物操纵和非经典生物操纵组合应用、以及经典生物操纵中不同鱼食性鱼类的组合放养对浮游动物群落结构影响的程度极其复杂,因此鱼类调控的力度以及调控力度与浮游动物群落结构上的变动之间的定量关系和因果关系是今后需要深入研究的科学问题.
本研究通过季节性监测山美水库浮游动物群落结构与水体理化因子,阐明了山美水库浮游动物群落密度主要表现为春夏季高,秋冬季低,入库区域高于其他区域的时空特点,生物量主要表现为夏秋季高,春冬季低,入库区域高于其他区域的时空特点. 本研究分析了浮游动物群落与水环境因子的关系,提炼了电导率、水温、硝态氮、溶解氧、透明度和高锰酸盐指数等关键环境因子. 结合近几年山美水库鱼类放养和捕捞措施的调整,初步分析了鱼类群落调控与浮游动物群落改变的关系,并进一步提出了相应的水质改善措施,可为湖泊、水库水环境保护和资源高效利用提供科学依据.
4 附录
附表Ⅰ和Ⅱ见电子版(DOI: 10.18307/2022.0619).
