不同林龄杉木根际与非根际土壤微生物群落特征
2022-11-09高李文彭紫薇韦明倩王玉哲胡亚林
郭 文, 高李文, 彭紫薇, 韦明倩, 王玉哲, 胡亚林, 刘 先
(福建农林大学 林学院, 福州 350002)
土壤微生物是土壤生态系统的重要组成部分,其参与有机质分解、养分循环和能量流动等生态过程,在维护生态系统的养分平衡中起着重要的作用[1-2]。土壤微生物不仅是土壤有效养分转化的动力,并且其本身也是土壤有效养分的供源和储源[3],其数量和组成对土壤环境变化具有高敏感性,能够对土壤环境变化快速做出改变,可以综合反映土壤质量和肥力在不同时间及不同环境内的变化情况[4-5]。同时,土壤微生物群落结构还可以反映土壤生态系统的稳定性,能够很好地指示在人工林经营管理过程中土壤生态功能的变化[6]。
土壤微生物群落受植被类型[7]、气候[8]、土壤环境状况等[9]因子的影响,这些因子相互作用共同影响土壤微生物数量和群落组成。同时,在人工林林分发育过程中,林内凋落物输入量、根系活力和微气候随林龄变化发生显著变化,从而使土壤微生物数量和群落组成在不同林龄间产生较大差异[10]。根际作为植物与外界进行物质和能量交换的主要场所,是土壤微生物活动最频繁的区域[11-12]。植物根系通过分泌各类有机物质来影响根际环境内的土壤性质[13],从而影响微生物在根际土壤中的数量和组成。使得不同林龄林木间根际和非根际土壤间微生物含量及组成产生较大差异。根际和非根际土壤微生物群落的差异使根际有效养分含量发生变化,直接影响林木的生长。因此,探究不同林龄根际和非根际土壤微生物含量及组成对了解林木生长发育过程中土壤质量变化具有重要意义。
杉木(Cunninghamialanceolata)是我国南方重要的速生用材树种,种植面积广泛,在我国南方林业生产中占有重要的地位[14],然而,随着杉木人工林面积的增加及长期的纯林经营及连栽等不良的经营措施,杉木人工林出现土壤理化性质恶化、养分循环失调、土壤地力衰退等问题[15-16],已经严重制约了杉木人工林的可持续性经营。磷酸脂肪酸(PLFA)在自然生理条件下相对恒定,许多PLFA可以作为微生物群落结构的生物标记物[17]。本文采用PLFA技术分析不同林龄杉木根际非根际土壤微生物群落特征,探究土壤微生物群落随林龄的变化特征和影响因子,以及不同林龄根际和非根际土壤微生物群落的差异。以期为中亚热带杉木人工林可持续性经营和维持其土壤肥力提供科学依据,以实现更大的效益和价值。
1 材料与方法
1.1 研究区与样地概况
试验区位于福建省中北部福建农林大学西芹教学林场,地理位置大致为北纬26°34′15″,东经118°05′41″,地处南方低山丘陵区,海拔180~200 m,坡度为25°左右。属中亚热带季风气候,年均温为17.3℃,年降水量为1 817 mm。本试验区土壤为黄红壤,土体含有少许石砾。主要林下植被主要有粗叶榕(FicushirtaVahl)、半边旗(PterissemipinnataL)、狗脊蕨(Woodwardiajaponica)、铁芒萁(Dicranopterislinearis)、海金沙(Lygodiumjaponicum)、荩草(Arthraxonhispidus)等。
在研究区内选取位置相近,立地条件基本一致的3个不同林龄(7,24,34 a)杉木人工林作为研究对象。在每个林龄杉木人工林分别设置4个20 m×20 m的样方,各样地间隔大于50 m,对样地基本情况进行调查,杉木不同林龄人工林样地基本概况见表1。

表1 不同林龄杉木人工林基本概况
1.2 土壤采样与处理
2019年8月,根据样地基本情况,在每块样地选取3棵生长状况一致的杉木作为标准木,采集根际和非根际土壤样品。根际土壤采用Riley等[18]的抖落法收集,先去除标准木地表凋落物及残留覆盖物,在基部顺着根系挖取细根,去除块不含根系的土壤,抖落粘附在根系表面的土壤作为根际土(Rhizosphere,R),混匀并装入自封袋内,共12个根际土壤样品;同时在标准木树冠投影范围外,沿S型选取5个取样点采集0—20 cm土壤样品作为非根际土(Non-rhizosphere,S),将各取样点土壤混匀,共12个非根际土壤样品,将采集后的土壤样品密封低温保存并迅速带回实验室。采集的土壤样品过2 mm筛,去除石块、根系以及土壤动植物,一部分土壤自然风干,研磨后用于土壤理化性质分析;另一部分置于-80℃冰箱中保存用于土壤微生物PLFA测定。
1.3 土壤理化性质和微生物PLFA分析

土壤微生物群落测定根据White等[20]的方法,将3 g冷冻干燥土壤加入土壤提取液(柠檬酸缓冲液∶甲醇∶氯仿=0.8∶1∶2)中浸提,用氮气吹干后溶于氯仿并利用SPE-Si柱进行纯化,纯化后加入甲醇甲苯混合液和KOH-甲醇溶液甲酯化,氮气吹干后用正己烷作为内标,上机用GC-MS分析仪进行分析,测定各磷脂脂肪酸含量,根据表2生物标记物表征各微生物群落。

表2 检测土壤微生物群落磷脂脂肪酸标记物
1.4 数据处理
采用SPSS 22.0进行双因素方差分析,来检验林龄、根际及其交互作用对土壤理化性质和微生物群落特征的影响,事后检验采用最小显著性差异法(Least-significant difference,LSD)法,考虑到森林土壤的异质性和重复样品数,显著性水平设为p<0.1。土壤微生物群落特征与土壤理化性质和养分的相关关系用Pearson相关分析检验。运用多元统计分析软件Canoco 5.0对不同林龄土壤微生物特征与土壤环境因子的关系进行冗余分析(Redundancy analysis,RDA)。
2 结果与分析
2.1 不同林龄杉木人工林根际和非根际土壤理化性质
土壤养分含量在不同林龄间具有显著差异(表3,p<0.1),随林龄增加,根际和非根际土壤全磷,可溶性有机碳含量(DOC)显著降低,在34 a最小,硝态氮含量随林龄增加显著增加,在24 a最大。铵态氮含量在34 a非根际土壤高于根际土壤,可溶性有机氮含量在7 a根际土壤高于非根际土壤。而其他养分含量在不同林龄及根际和非根际土壤间均无显著差异。根际土壤pH值随林龄增加显著降低,在非根际土壤中则先减后增;根际和非根际土壤含水率随林龄增加均先增后减,在24 a达到最大。

表3 土壤基本理化性质
2.2 不同林龄杉木林土壤各类群微生物PLFAs含量特征
不同林龄杉木林根际和非根际土壤微生物PLFAs含量进行分析结果表明,无论在根际还是非根际土壤,细菌群落PLFA含量均为最高,丛枝菌根真菌PLFA含量为最低(图1)。根际土壤各类微生物群落PLFAs含量在不同林龄间存在差异,均在34 a含量最高,7 a含量最低。根际土壤总PLFAs含量表现为34 a>24 a>7 a。而在非根际土壤中,细菌和放线菌PLFAs含量在7 a最高,在34 a最低,而真菌则在24 a最高。根际非根际土壤间,细菌PLFAs含量在34 a根际土壤显著高于非根际土壤,但在7 a和24 a两个人工林间没有显著差异,(表4,p<0.1)。其他微生物群落在根际非根际间均没有显著差异。
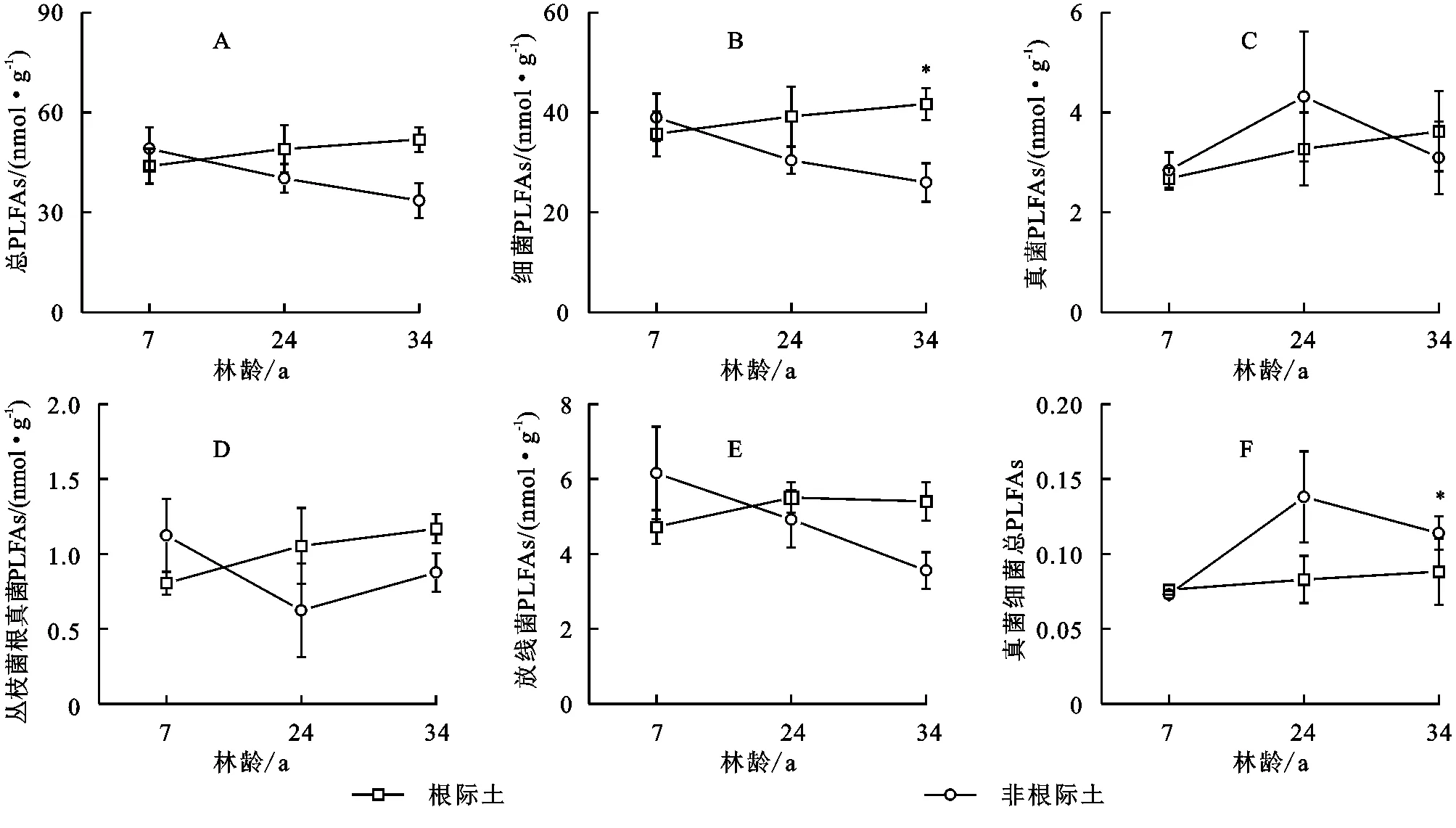
注:*表示相同林龄根际非根际土壤差异性显著,下图同。图1 不同林龄杉木林土壤各类群微生物PLFAs含量

表4 双因素方差分析林龄、根际及两者交互作用对土壤各类群微生物PLFAs的影响
随林龄增加,根际土壤革兰氏阳性菌革兰氏阳性菌(G+)、革兰氏阴性菌(G-)、支链饱和脂肪酸(TBSAT)、单烯不饱和脂肪酸(MONO)、环丙烷脂肪酸(Cy)PLFAs含量均在34 a最高,7 a最低,而饱和脂肪酸(SAT)PLFAs含量在7 a最高,24 a最低。非根际土壤细菌群落各类PLFAs含量均在34 a达到最低(图2)。在7 a和24 a杉木人工林中,根际土壤和非根际土壤细菌群落各类PLFAs含量没有显著差异,但在34 a中,根际土壤革兰氏阴性菌(G-)和环丙烷脂肪酸(Cy)显著高于非根际土壤。
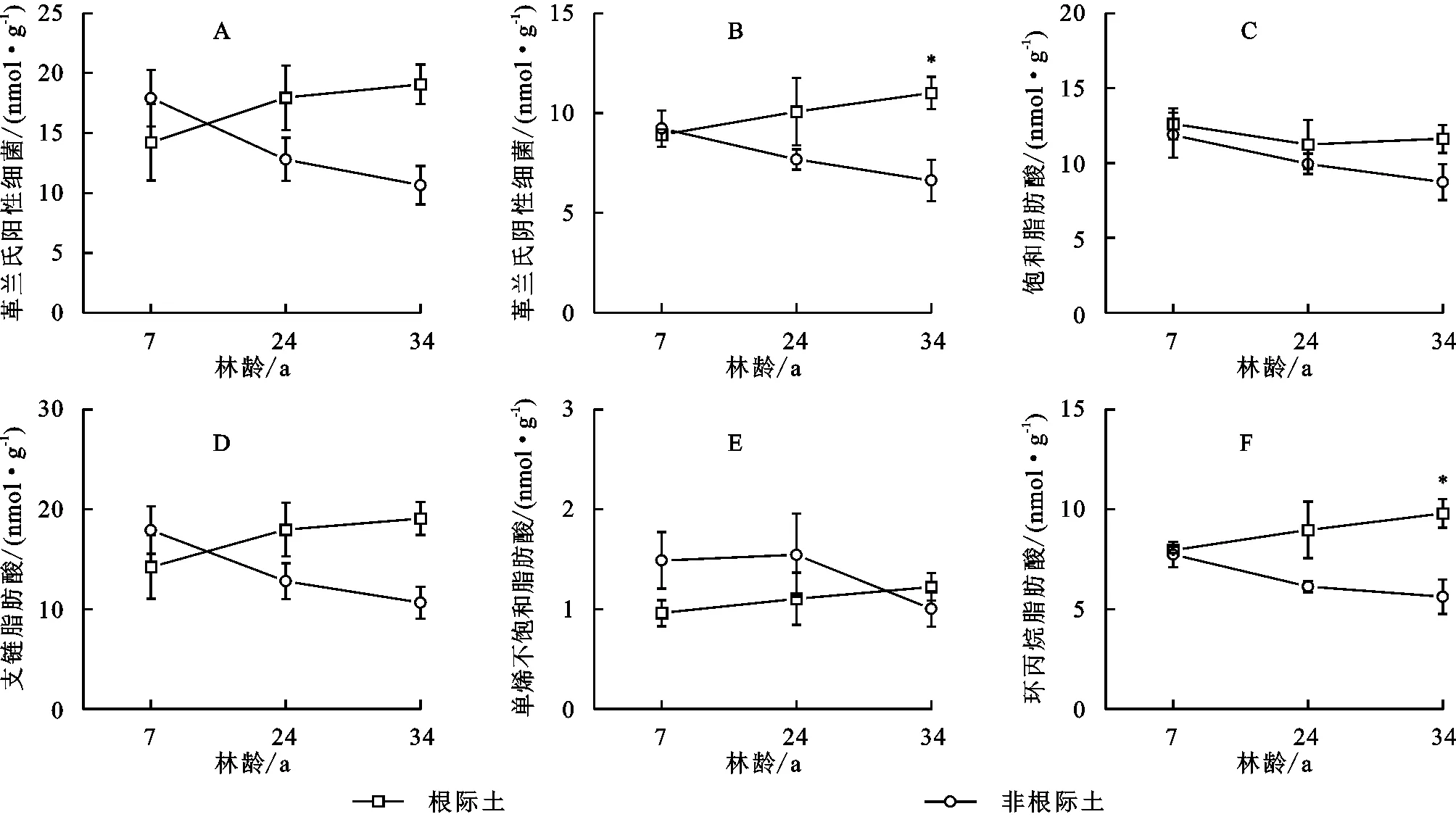
图2 不同林龄杉木林土壤细菌群落PLFA含量
2.3 不同林龄杉木林根际和非根际土壤微生物群落结构
对表征土壤微生物群落结构的真菌∶细菌、革兰氏阳性菌(G+)∶革兰氏阴性菌(G-)、环丙烷脂肪酸(Cy)∶单烯不饱和脂肪酸(MONO)、MONO∶支链饱和脂肪酸(TBSAT)进行分析,林龄对各微生物结构没有显著影响,根际和非根际土壤间Cy∶MONO和MONO∶TBSAT具有显著差异,林龄和根际的交互作用对MONO∶TBSAT具有显著影响(表5)。7 a杉木人工林中,Cy∶MONO根际显著高于非根际(图3),而34 a杉木人工林中,MONO∶TBSAT非根际土壤显著高于根际土壤,Cy∶MONO根际土壤显著高于非根际土壤。

表5 双因素方差分析林龄、根际及两者交互作用对微生物群落PLFAs结构的影响

图3 不同林龄杉木林土壤微生物群落结构
2.4 土壤理化性质对土壤微生物群落特征的影响
相关性分析表明,细菌、总PLFA量和G+与土壤有效磷含量呈极显著正相关(表6,p<0.05),与铵态氮含量呈极显著负相关;真菌与硝态氮含量呈显著正相关(p<0.1);丛枝菌根真菌与土壤碳磷比呈极显著正相关,与氮磷比呈显著正相关;放线菌与土壤有效磷含量呈显著正相关;G-与土壤有效磷含量和土壤可溶性有机氮含量呈极显著正相关;真菌/细菌与硝态氮含量呈显著正相关;G+∶G-与土壤有效磷含量呈极显著正相关(p<0.01),与铵态氮含量呈显著负相关。冗余分析(RDA)结果表明第一轴和第二轴分别解释了土壤微生物群落结构变异的59.4%,13.4%(图4)。土壤环境对微生物群落有显著影响,其中土壤有效磷、铵态氮、可溶性有机氮、全碳、全氮含量对土壤微生物群落结构的影响较大,尤其是土壤有效磷、铵态氮含量的影响最大。

表6 土壤微生物群落结构与土壤理化性质相关性分析

图4 不同林龄杉木土壤微生物结构与土壤因子之间的冗余分析
3 讨 论
在森林生态系统中,林木林龄的变化能够改变土壤环境,使土壤微生物随环境变化而发生改变[10]。本研究中,随着杉木林龄的增加,根际土壤各类微生物PLFAs含量不断增加,而在非根际土壤中不断减少,并且细菌含量在34 a根际土壤显著高于非根际土壤。这可能与杉木自身特性及土壤环境变化有关,在根际土壤中,杉木主要通过根系活动来改变根际环境,随着杉木林龄的增加,根系显著生长,根系活力和分泌速率增强[21],并且根际沉积使养分在根际富集[22],进而促进了根际微生物的生长。而非根际土壤直接与地表腐殖质层接触,其土壤微生物受凋落物及土壤养分含量的影响较大[23]。在杉木人工纯林的生长发育过程中,其凋落物较少,因而其对土壤养分具有高吸收,低归还的特点[24],而在其造林前期,采伐迹地保留了大量的枝叶,增加了养分归还量,同时炼山等措施产生的黑炭能够在短期内显著提高土壤中有效养分含量[25-26],使非根际土壤微生物含量在生长前期较低,随着养分的消耗,土壤微生物含量逐渐增加。在土壤微生物群落中,细菌群落占主导地位,已有研究表明,土壤pH值是影响土壤细菌群落组成的重要因素[27]。相对较低的pH值适合细菌的生长,使得细菌在根际土壤高于非根际土壤。
真菌和细菌是土壤生态系统中重要的微生物群落,其参与有机质的分解,是土壤肥力的重要指标之一,细菌占主导地位的土壤肥力较高[28],此外,真菌∶细菌能够表征生态系统的稳定性,比值越大,表明土壤系统越稳定[29]。本研究发现,细菌在杉木人工林土壤处于主导地位,且真菌:细菌的比值随林龄的增加没有显著差异,说明杉木人工林林龄的增加并没有改变土壤生态系统的稳定性。G+∶G-常用来指示土壤营养状况,比值越大营养胁迫越大[30],同时Cy∶MONO和MONO∶TBSAT也可以表征养分限制和环境胁迫的程度[29]。本研究中,根际和非根际土壤G+∶G-,Cy∶MONO和MONO∶TBSAT在不同林龄间没有显著差异,但Cy∶MONO和MONO∶TBSAT在34 a根际土壤均显著高于非根际土壤。这表明杉木人工林随林龄变化营养状况没有较大变化,但在34 a中,非根际土壤微生物受到的环境胁迫(如养分含量、水分等)大于根际土壤。
土壤微生物结构与群落组成受多种因素影响,一方面,其受植被类型、气候、人类活动等因素的影响,另一方面,还受到土壤养分含量的影响[31]。相关分析和冗余分析表明,土壤养分含量是土壤微生物群落结构和组成的主要影响因子,这与前人对土壤微生物含量的研究结果一致[32]。其中铵态氮与细菌、G-和总PLFAs含量呈显著负相关。已有研究表明,土壤中过量的铵态氮会抑制微生物吸收利用其他形态的氮源,从而抑制微生物的生长[33]。土壤有效磷与各微生物群落(除真菌外)呈显著正相关,在中亚热带森林中土壤普遍存在磷限制,当土壤存在磷限制状况时,土壤微生物能够分泌较多与磷相关的酶来提高有机磷的矿化,促进有效磷含量[34]。土壤真菌含量与硝态氮呈显著正相关,丛枝菌根真菌与C∶P和N∶P呈显著正相关,说明土壤中N和P含量一定程度上会影响土壤微生物群落。此外,可溶性有机物是土壤养分迁移及损失的主要形式,其主要来源于微生物的活动和有机质的分解[10],本研究中,土壤DON和DOC对微生物群落呈显著正相关关系,对微生物的活动具有显著影响。
4 结 论
随林龄的增加,杉木人工林根际土壤各类微生物数量不断增加,而在非根际土壤中不断减少,细菌数量在34 a根际土壤显著高于非根际土壤,在其他微生物及其他林龄间均无显著差异。
土壤有效磷和铵态氮含量是驱动土壤微生物变化的最主要因素,主要原因是中亚热带地区主要存在磷限制,土壤微生物能够分泌更多的有效磷供杉木吸收利用。因此,在杉木人工林经营管理过程中,应当适当增施磷肥,以提高土壤微生物数量,确保杉木的良好发育。
