不同养分添加对高寒植物功能性状的影响
2022-11-04叶琼丹李永慧张子杨李希来李兰平
叶琼丹, 任 飞, 李永慧,4, 张子杨, 樊 娜, 冶 俊, 李希来, 李兰平*
(1. 青海大学省部共建三江源生态与高原农牧业国家重点实验室, 青海 西宁 810016; 2. 青海三江源草地生态系统国家野外科学 观测研究站, 青海 西宁 810016; 3. 青海大学农牧学院, 青海 西宁 810016; 4. 青海大学生态环境工程学院, 青海 西宁 810016)
植物功能性状(Plant functional trait,PFT)是可确定植物如何响应环境因素、影响其余营养级和生态系统功能的一些植物形态结构、生理生化和物候特征[1-2];作为植物联系环境的纽带,在全球变化、古植被恢复、生态保护和恢复等领域得到了广泛的关注,是目前生态学研究的热点之一[3]。关于植物功能性状对全球变化因子的响应,目前的研究更多关注于气候变化和CO2浓度等因子,而大气氮、磷沉降对植物功能属性的影响尚缺乏更深入的研究[4-6]。
氮(Nitrogen,N)、磷(Phosphorus,P)是植物生长必需的两种营养元素。传统观点认为,受成土和气候因子的影响,植物的生长在高纬地区通常受氮限制,在低纬地区则受磷限制[7]。也有研究表明,植物生长会同时受到氮、磷限制,但氮磷共同限制的程度因氮、磷交互作用程度与方向的不同而相异[8]。在全球环境变化背景下,大气氮、磷沉降逐年增加,额外的氮、磷输入将对生态系统的结构和功能产生深远影响[9-10]。例如,詹书侠等在内蒙古草原基于羊草对氮、磷添加的响应实验发现,氮添加会显著提高羊草地上生物量和总生物量,而磷添加显著提高了羊草地下生物量[11];齐瑞等研究表明氮、磷混合添加会加速植物群落生物量的增加和物种丰富度的降低[12];同时,周一平等的研究发现氮添加有利于提高植物对光的竞争,磷添加可以提高从属种的竞争优势,从而改变群落结构和功能[13]。总之,氮、磷养分元素与植物功能性状的关系较为复杂,也表明植物可通过功能性状的可塑性去适应氮、磷养分变化的环境[4,14]。
目前,针对高寒地区不同梯度养分单独添加和混合添加对植物功能性状的影响尚未见系统性研究报道,氮、磷添加对植物功能性状的交互作用影响尚不明确。因此,在大气氮、磷沉降逐年加剧的背景下有必要从植物功能性状对氮、磷养分添加的响应角度去探讨高寒植物对全球变化的适应机制。为此,本研究依托中国科学院海北高寒草地生态系统国家野外科学观测研究站(简称海北站)的多梯度氮磷添加实验平台,通过分析10种常见高寒植物的高度(Height,H)、叶面积(Leaf area,LA)、比叶面积(Specific leaf area,SLA)、叶干物质含量(Leaf dry matter content,LDMC)、茎密度(Stem tissue mass density,STD)、地上生物量(Individual aboveground biomass,IAGB)随不同梯度氮、磷单独添加及混合添加的变化,研究高寒植物功能性状对不同养分添加的响应,为预测大气氮、磷沉降加剧背景下高寒植物功能性状的演变趋势提供依据。
1 材料与方法
1.1 研究区概况
研究地点位于中国科学院青海海北高寒草地生态系统国家野外科学观测研究站(简称海北站)附近,海北站地处青藏高原东北隅的青海海北藏族自治州门源回族自治县境内,样地中心坐标37°39′15.407″ N,101°19′54.200″ E,海拔3 231 m,为典型的高寒草甸[13]。研究区以山间滩地和丘陵低山为主,属高原大陆性气候,无明显的四季之分,仅有冷暖两季之别;年平均气温—1.7℃,最冷月(1月)平均气温—15.2℃,最热月(7月)平均气温9.9℃[15-16]。该区干湿季节分明,雨热同期,年降水量范围425.36~850.4 mm之间,平均为582.1 mm,降水80%分布于植物生长季的5—9月,20%分布于冷季,平均年日照时间2 462.7 h[17]。站区土壤发育年轻,为草毡寒冻雏形土(高山草甸土)。样地植被类型为以矮生嵩草(Kobresiahumilis)为建群种的高寒嵩草草甸,优势种为垂穗披碱草(Elymusnutans)、异针茅(Stipaaliena)、山地早熟禾(Poaversicolor)等,常见种包括甘肃棘豆、紫羊茅(Festucarubra)、美丽风毛菊、棉毛茛(Ranunculusmembranaceus)、麻花艽(Gentianastraminea)等,群落内大多数植物为多年生草本,但稀有种包含一种灌木,为矮生忍冬(Loniceraminuta)[13]。
1.2 试验设计
试验采取完全随机区组设计(表1),其中N添加设置3个水平,分别为N0(无养分添加)、低水平N100(10 g·m-2·a-1)和高水平N200(20 g·m-2·a-1),P添加设置3个水平,分别为P0(无养分添加)、低水平P50(5 g·m-2·a-1),和高水平P100(10 g·m-2·a-1),共9个处理,每个处理设置4个重复(区组,Block),共计36个小区(Plot),区组和小区之间均设置1 m的缓冲带(过道)。实验样地于2017年建立围栏,2018年开始养分添加处理,其中氮以尿素(Urea)的形式添加,磷以重过磷酸钙(Triple superphosphate)的形式添加。施肥方式是在6月的第一周和第二周分两次将预先称好的颗粒状肥料均匀撒施于各小区内。选择阴天下雨之前或傍晚(晴天时)施肥,以便雨水或次日形成的露水将肥料迅速溶解进入土壤。在实验期间(每年4—10月)对样地进行围栏封育管理,实验结束后短期放牧藏系绵羊1—2个月以去除地表枯落物,并在返青期前人工清除绵羊排泄物和剩余的枯落物。
表1 完全随机区组设计表Table 1 Completely randomized block design table
本研究以实验样地植物群落中的10种常见物种为研究对象,分别为垂穗披碱草、圆萼刺参(Morinachinensis)、钝裂银莲花(Anemoneobtusiloba)、美丽风毛菊、甘肃棘豆、钉柱委陵菜(Potentillasaundersiana)、麻花艽、钝苞雪莲(Saussureanigrescens)、棉毛茛、重冠紫菀(Asterdiplostephioides)。
1.3 取样和测定方法
2021年6—8月,在小区内随机选取5~30株处于盛花期的10种代表性物种(垂穗披碱草30株,钝裂银莲花20株,棉毛茛20株,其余物种均5株),每株沿基部剪取地上部分后分别装入样方号及重复号不同的密封袋中以减少其水分散失,避免叶片萎缩。取样完成后迅速将植株样品带回室内进行性状测量,在室内记录每株的高度、叶面积、茎的直径等。
株高(H)测定:用卷尺测定每株单株的基部到冠层顶端的长度。
茎干重(Stem dry weight,SDW)测定:在实验室内将茎叶分离,将每株单株截取离地面1 cm处取下5 cm(L)的茎节,用电子游标卡尺量取垂直切面的长(a)、短(b)直径,用烘箱在105℃下杀青0.5 h后在65℃下烘干至恒重,后用万分之一天平称重。
地上生物量(IAGB)测定:将处理后的植株分别装入已编号的信封中,用烘箱烘干至恒重后称重。
叶片饱和鲜重(Leaf fresh weight,LFW)测定:将叶片从密封袋中取出,迅速用吸水纸吸去叶片表面的水分,用万分之一天平称取特定叶片鲜重。
叶面积(LA)测定:由扫描仪扫描随机选定成熟完整的叶片后使用Image J软件计算得出。
叶片干重测定(Leaf dry weight,LDW):将测定完叶面积的叶片装入已编号的信封中,与上同,烘干至恒重后称重。
PFT的计算公式如下:
(1)鲜茎体积(Volume,V)
(2)比叶面积(SLA)
(3)叶干物质含量(LDMC)
(4)茎密度(STD)
1.4 数据分析
采用单因素方差分析和最小显著差异(LSD)多重比较方法,分析不同养分添加处理对总PFT和各物种PFT的影响。利用双因素方差分析,将不同水平的氮、磷添加处理作为固定因子,检验氮、磷添加对PFT是否有交互作用影响。由于各物种样本采集数量不同,以上分析采用了各物种功能性状在各小区的平均值。所有统计分析利用SPSS 26完成,Origin 2021绘图。
2 结果与分析
2.1 PFT对氮、磷添加的总体响应
单因素方差分析结果表明氮单独添加对6种功能性状指标均无显著影响;磷单独添加对SLA有显著影响(P<0.05),对H,LA,STD,IAGB均无显著影响,仅高磷单独添加对LDMC有显著影响(P<0.05);氮、磷混合添加(N100P50,N100P100与N200P50,N200P100)均显著增加了植物的H和SLA(P<0.05),显著降低了植物的LDMC和STD(P<0.05),仅高氮混合添加显著增加植物LA(P<0.05),所有氮、磷混合添加条件对IAGB均无显著影响(图1)。氮、磷混合添加下,H表现为N200P100>N100P100>N200P50>N100P50,H在高氮高磷的条件下增加的幅度最大,与对照相比增加了46.53%;混合添加下LA,SLA,STD和LDMC变化幅度表现为N200P50>N200P100>N100P100>N100P50,均在高氮低磷的条件下变化的幅度最大,其LA和SLA比对照多85.33%和34.09%,STD和LDMC比对照少20.39%和24.84%(图1)。双因素方差分析结果表明氮添加与磷添加对6种PTF均无显著交互作用影响(表2)。
2.2 不同物种PFT对氮、磷添加的响应
氮单独添加对圆萼刺参大部分性状指标有显著影响(P<0.05),对钝裂银莲花、钉柱委陵菜、麻花艽、美丽风毛菊、重冠紫菀和钝苞雪莲大部分性状指标无显著影响,对棉毛茛和甘肃棘豆6种性状指标均无显著影响;高氮单独添加对垂穗披碱草大部分性状指标有显著影响(P<0.05)。H,LA,SLA,IAGB随着氮添加量的增加总体呈增加的趋势,LDMC和STD出现下降的趋势。在高氮单独添加条件下,垂穗披碱草H、麻花艽LA和IAGB以及钝苞雪莲SLA增加的幅度最大,钉柱委陵菜LDMC和垂穗披碱草STD减少的幅度最大(表3)。

图1 氮、磷单独添加及混合添加对植物功能性状的影响Fig.1 Effects of nitrogen and phosphorus addition alone and mixedly on plant functional traits注:不同小写字母表示不同氮、磷混合添加处理间差异显著(P < 0.05),ns表示无显著差异。N0P0表示对照;N100,N200表示低、高氮水平;P50,P100表示低、高磷水平Note:Different lowercase letters indicated that there were significant differences between different nutrient addition treatments at the 0.05 level,ns indicates no significant difference. N0P0 represents control;N100,N200 represents low and high N levels;P50,P100 represents low and high P levels

表2 氮、磷添加及其交互作用对植物功能性状影响Table 2 Effects of nitrogen,phosphorus addition and their interaction on plant functional traits
磷单独添加对钝裂银莲花大部分性状指标有显著影响(P<0.05),对棉毛茛、垂穗披碱草部分性状指标有显著影响(P<0.05),对圆萼刺参、麻花艽、钉柱委陵菜、重冠紫菀和钝苞雪莲大部分性状指标无显著影响,对甘肃棘豆、美丽风毛菊6种性状指标均无显著影响。钝裂银莲花LA、垂穗披碱草IAGB和重冠紫菀SLA在高磷处理下增幅最大,钉柱委陵菜LDMC和STD减幅最大,垂穗披碱草H在低磷条件下增加的幅度比高磷大(表3)。
氮、磷混合添加对除甘肃棘豆和美丽风毛菊外其余8种植物大部分性状指标有显著影响(P<0.05,表3)。其中对圆萼刺参、麻花艽IAGB,垂穗披碱草SLA,钝苞雪莲LA和IAGB无显著影响,对甘肃棘豆和美丽风毛菊H有显著影响(P<0.05,表3)。在不同浓度梯度氮、磷养分添加处理中,不同物种间的PFT对养分添加的响应整体幅度为NP>P>N。

表3 氮、磷单独添加及混合添加对10种植物功能性状的影响Table 3 Effects of nitrogen and phosphorus addition alone and mixedly on functional traits of 10 species
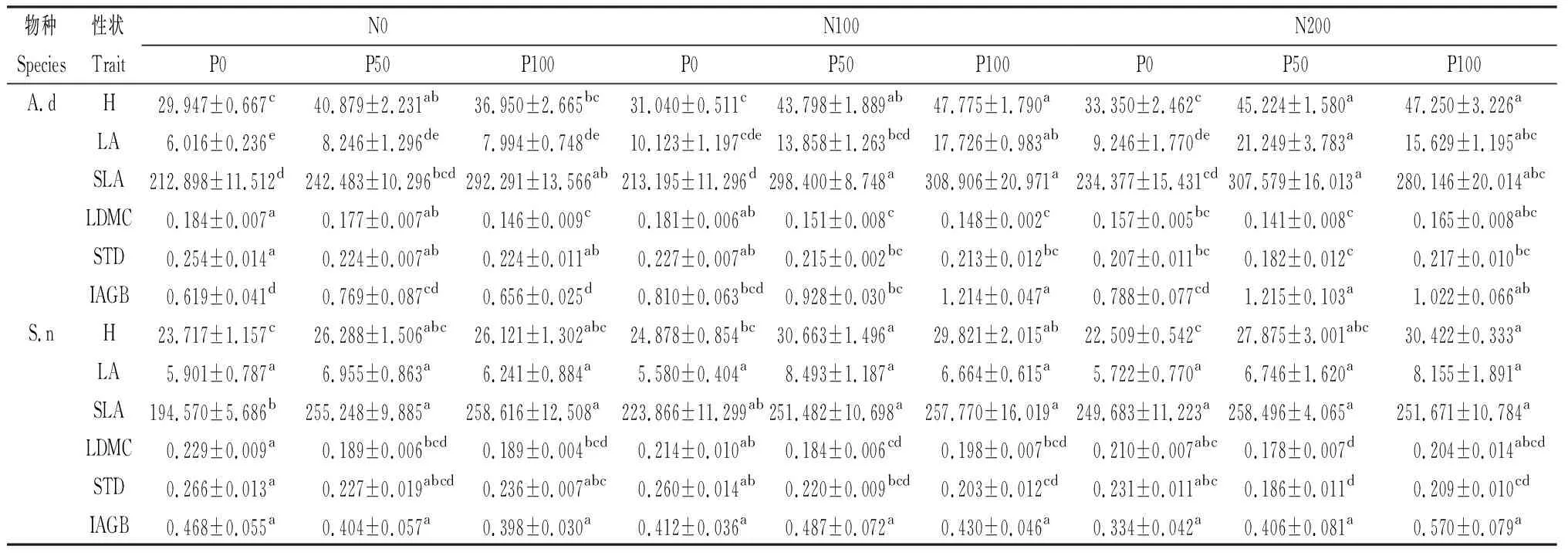
续表3
3 讨论
3.1 氮、磷添加对PFT的影响
本研究发现,氮、磷单独添加对研究区域植物功能性状影响较弱,仅磷单独添加对SLA和LDMC有显著影响;氮、磷混合添加对除了IAGB之外的5种功能性状均有显著影响,但氮添加与磷添加对6种PTF均未表现出显著交互作用影响。这说明研究区域植物的生长可能受到了氮磷共同限制的影响,但是土壤中植物可利用的氮和磷尚未缺乏到影响另一养分元素的吸收和利用的程度。
植物H仅在氮、磷混合添加下显著增加。该结果说明研究区域植物H主要受到氮、磷共同限制的影响,在解除氮、磷的限制后植物可通过提升高度去捕获更多光能以提高对光的竞争。此结果与万宏伟等[18]施肥实验研究结论相似,验证了在限制性土壤养分得到满足后,植物会降低对养分竞争的强度,通过利用功能性状的可塑性来提高对光资源的竞争[19-20]。
在植物功能性状特征中,LA,SLA,LDMC和STD能反映植物获取资源的能力[21-22]。本研究发现,LA,SLA,LDMC和STD均在高氮低磷的条件下变化的幅度最大,即在磷添加量达到一定程度时LA,SLA不再增加,STD,LDMC不再减少。这说明,随着土壤中磷可利用性的增加,植物会采取获取型策略,增加光合作用的能力,较快的获取养分资源以适应环境[19]。但当氮、磷混合添加的养分达到一定浓度时,植物会从获取型策略转换为保守型资源利用策略,减小LA,SLA,LDMC和STD的变化幅度,减缓养分的循环利用以适应高浓度的养分环境[23]。
本研究发现,IAGB总体上不受氮、磷添加及其交互作用的显著影响。该结果与部分学者发现氮添加会促进羊草生长,提高羊草群落的地上生物量和个体生物量的结果不一致[11,24]。这可能是因为在本实验中虽然氮、磷混合添加使得植物H升高,但STD,LDMC减小,从而使IAGB没有显著差异。同时各物种IAGB在氮、磷添加下变化趋势不一致,如甘肃棘豆与钝苞雪莲IAGB均在部分养分添加处理下出现了下降的情况,这也可能是导致IAGB总体上在养分添加处理下没有显著差异的原因。
3.2 氮、磷添加对不同物种PFT的影响
本研究发现,10种物种的PFT对氮、磷单独添加的响应具有一定的物种差异性。其中,圆萼刺参PFT对氮单独添加敏感,垂穗披碱草PFT在高氮单独添加下大部分性状指标有显著变化,其余物种大部分或全部PFT在氮单独添加下无显著变化;钝裂银莲花对单独添加磷敏感,除了钝裂银莲花、棉毛茛和垂穗披碱草外7种物种大部分或全部PFT在高磷单独添加下无显著变化。氮、磷混合添加对除美丽风毛菊和甘肃棘豆外其余8种植物大部分性状指标均有显著影响。该结果说明研究区域大部分植物的生长可能主要受到氮磷共同限制的影响,虽然部分物种如甘肃棘豆、美丽风毛菊等的PFT对于养分添加的响应表现出了较大的差异性,但大部分物种PFT对于养分添加的响应是一致的。这可能是因为虽然植物对于环境变化的响应受到了遗传背景的限制,但是在环境选择压力下功能性状会倾向于趋于一致[25-26]。因此,高寒植物不同物种对于全球变化背景下氮磷养分供给量的改变存在不同的适应策略,但其功能性状总体上会有相对确定的演变趋势。
与对照相比,垂穗披碱草H在高氮高磷添加时增加幅度最大,麻花艽LA、棉毛茛SLA、重冠紫菀IAGB、钉柱委陵菜LDMC和棉毛茛STD在高氮低磷添加时变化幅度最大。这说明随着养分含量的增加,垂穗披碱草作为优势种,为维持在群落中的稳定地位,通过不断提高对高氮高磷养分的吸收和转化,增加H以占据更高的生境空间,获得更丰富的光资源,达到对下层物种进行优势压制的目的[27]。此结果与王萌等[28]研究克氏针茅草地植物叶片功能性状对氮添加的响应的结果相似。麻花艽、棉毛茛、钉柱委陵菜、重冠紫菀位于群落下层,在特定养分条件得到满足后,麻花艽、棉毛茛会通过增大叶片大小和茎秆机械支撑的强度来增加对光照的截取能力[29-30]。钉柱委陵菜、重冠紫菀主要通过减少LDMC和增加IAGB的方式来转换资源利用策略,增强光合作用能力,较快的获取养分资源,提升种间竞争优势[31-32]。这与杨晓霞等[33]研究青藏高原高寒草甸植物群落生物量对氮、磷添加的响应的结果较为一致。
4 结论
本研究表明,PFT主要受到氮、磷混合添加的影响,氮、磷单独添加对PFT的影响较弱。在物种水平上,大部分物种主要受氮、磷混合添加的影响,对氮、磷单独添加不敏感。根据本研究结果,我们预测在大气氮、磷沉降日益加剧的背景下,高寒植物未来会倾向于更高、叶片更大更薄,但茎密度会减小,植株个体地上生物量变化不明显或仅略微增加。因此,大气氮、磷沉降加剧可能通过推动高寒植物功能性状的演变对高寒草地生态系统功能产生影响,但该影响尚待研究和评估。此外,相关研究还需考虑更多维度功能性状对不同养分添加响应,以更全面地了解高寒植物对不同养分添加的适应策略。
