外源激素对光皮桦种子萌发中抗氧化酶活性的影响
2022-10-27韦虹日苗艺明刘世男
韦虹日,苗艺明,刘世男,杨 梅*
(1.广西雅长林场,广西 百色 533000;2.广西大学 林学院,南宁 530004)
光皮桦(Betulaluminifera)是桦木科桦木属落叶乔木[1],主要分布在秦岭以南以及两广地区,生长快、耐瘠薄,材质优良、用途广,是南方主要速生树种之一[2],具有很高的经济价值。光皮桦繁育技术研究对丰富南方优良造林树种苗木,促进缓解全国木材资源短缺有重要作用。
林木种子预处理是播种育苗的关键技术环节之一,也是培育良种壮苗的第一步[3]。外源激素作为具有生理调节功能的植物生长调节剂,通过外源激素的质量浓度与配比变化,调控种子萌发过程中的SOD、POD和CAT等一系列抗氧化酶活性[4],从而调节种子体内代谢及萌发进程,实现林木不同生长发育阶段的需求[5]。李婉茹[6]研究发现赤霉素处理利于紫斑牡丹(Paeoniarockii)种子POD活性提升,促进种子萌发。赖华燕等[7]发现高浓度赤霉素和低浓度脱落酸对提高福建青冈(Cyclobalanopsischungii)种子萌发率和种子内POD、SOD、CAT酶活性作用显著。陶汉之[8]等研究发现赤霉素处理可以调高茶籽萌发过程中的CAT酶活性。余春和[9]等研究表明一定浓度GA3、IAA、IBA能促进光皮桦种子萌发,但生理机理尚不清楚。为提高光皮桦播种育苗种子发芽率和苗木质量,给制定播种育苗技术提供依据,我们研究了不同浓度GA3、IAA和IBA处理下光皮桦种子萌发过程中的抗氧化酶含量变化,分析了这3种激素促进种子萌发的机理。
1 材料与方法
1.1 试验材料
试验所用种子采自于广西雅长林场生长正常、健壮,中龄期母树的成熟饱满健康种子,置于4 ℃备用。
1.2 试验设计与布置
选择赤霉素(GA3)、吲哚乙酸(IAA)、吲哚丁酸(IBA)等3种外源激素对光皮桦种子浸种处理24 h,均设置10、30、50 mg·L-1等3个浓度梯度,以蒸馏水浸泡种子作为对照;每个处理设置6个重复,每个重复100粒种子。
外源激素处理前,选取干净光皮桦种子浸入0.1%高锰酸钾溶液10 min进行表面消毒,蒸馏水冲洗多次至高锰酸钾味道消失。种子激素处理后放置在铺有单层滤纸的无菌培养皿中,每日定时定量补充蒸馏水并观察统计种子萌发状况。
取干燥种子(M)、未萌发种子(M0,浸泡24 h后)、露白种子(M1)、胚根长度与种子等长种子(M2)、苗期种子(M3)等5个时期的种子样品,用于生理指标测定。
1.3 抗氧化酶活性和MDA含量测定
取光皮桦种子样品0.25 g,加入浓度0.05 mol·L-1、pH值=7的磷酸缓冲液10 mL及少许石英砂,在冰浴中研磨成匀浆,再分多次加入10 mL缓冲液冲洗并全部转移至50 mL离心管中,摇匀,离心20 min(4 ℃,5 000 rpm),取上清液测定抗氧化酶活性。
测定MDA含量采用硫代巴比妥算法,测定POD活性采用愈创木酚法[10],测定SOD活性采用氮蓝四唑光化还原法[11],测定CAT活性采用紫外吸收法[10],利用多功能酶标仪测定样品吸光度值。
1.4 数据分析
采用Excel 2010完成所有数据的整理与作图,采用SPSS 21.0统计分析软件对种子酶活性指标进行显著性分析。
2 结果与分析
2.1 不同浓度GA3、IAA、IBA浸种对抗氧化酶活性的影响
2.1.1 赤霉素 如图1所示,经GA3处理后的种子在萌发过程中,体内POD、CAT、SOD活性随GA3浓度升高而增强,与对照处理组差异显著(P<0.05),萌发过程中种子内MDA含量高于对照组且存在显著差异(P<0.05)。
在萌发过程M0至M3的4个时期中,低浓度GA3(10 mg·L-1)处理后的光皮桦种子内POD活性较对照组降低,分别降低9.8%、14.7%、20.0%、15.6%。而中高浓度GA3(30 mg·L-1、50mg·L-1)处理后的种子萌发过程中体内的POD酶活性明显高于对照组与低浓度处理组(10 mg·L-1)(P<0.05,图1b),且随GA3浓度的增加,POD酶活性也随之增强。高浓度GA3(50 mg·L-1)处理后,种子体内POD酶活性较对照组分别升高30.8%、21.9%、27.7%、27.7%。
GA3浸种处理后的光皮桦种子体内CAT、SOD酶活性显著高于对照组(P<0.05,图1c、图1d),并且随GA3浓度升高而增强。在M0至M3的4个时期内,高浓度GA3(50 mg·L-1)处理后,种子体内CAT活性较对照组分别增高39.1%、35.6%、31.1%、40.3%(图1c),SOD活性较对照组分别增高42.6%、51.4%、47.4%、56.3%(图1d)。
使用GA3处理后种子体内的MDA含量呈先增高再降低的变化趋势(图1a)。在M1时期,GA3浓度为10 mg·L-1时,种子体内的MDA含量达到峰值,较对照处理组提高79.2%;在M0、M2、M3等3个萌发阶段中,当GA3浓度为50 mg·L-1时,种子体内MDA含量出现最高值,分别较对照提高60.2%、178.5%、46.8%。

图1 不同浓度赤霉素处理下的种子抗氧化酶活性
2.1.2 吲哚乙酸 光皮桦种子体内POD酶活性随IAA浓度增加,整体呈现先升高后下降的趋势(图2b)。光皮桦种子经低中浓度IAA(10、30mg·L-1)处理后,POD活性增强,并随IAA浓度升高而增高(图2b)。高浓度IAA(50 mg·L-1)处理后,M0至M3的4个不同时期的种子体内POD活性均降低,较对照组分别降低19.4%、15.4%、18.3%、19.3%。在M0至M3的4个萌发阶段中,当IAA质量浓度为30 mg·L-1时,POD活性达到最高,此时POD活性分别较对照组增高27.0%、25.1%、19.9%。
整个萌发过程中,光皮桦种子体内CAT、SOD活性随IAA浓度增加分别呈现先升高再下降的变化(图2c、图2d)。在M0至M3的4个时期中,使用30 mg·L-1质量浓度IAA处理后,种子体内的CAT、SOD活性达到最高值,CAT活性较对照组提高35.1%、13.8%、19.9%,SOD活性较对照组提高49.6%、41.4%、34.5%、32.6%。
随IAA浓度的增加,萌发过程中种子体内的MDA含量逐步增高(图2a)。同一浓度处理下,M3时期种子MDA含量高于其他时期。经IAA处理后的光皮桦种子在M3时期的MDA含量较M0时期分别增高43.3%、43.3%、42.0%、43.9%。

图2 不同浓度吲哚乙酸处理下的种子抗氧化酶活性
2.1.3 吲哚丁酸 光皮桦种子经低中浓度IBA(10、30 mg·L-1)浸泡处理后,种子体内POD、CAT、SOD酶活性均增高,显著高于对照处理组(P<0.05),且抗氧化酶活性随IBA浓度的增加而增高;经高浓度IBA(50 mg·L-1)浸种处理后,光皮桦种子体内POD、CAT、SOD酶活性降低。光皮桦种子萌发过程中,POD、CAT、SOD活性随着IBA浓度的增加整体呈现先增高后下降的变化趋势(图3b、图3c、图3d)。
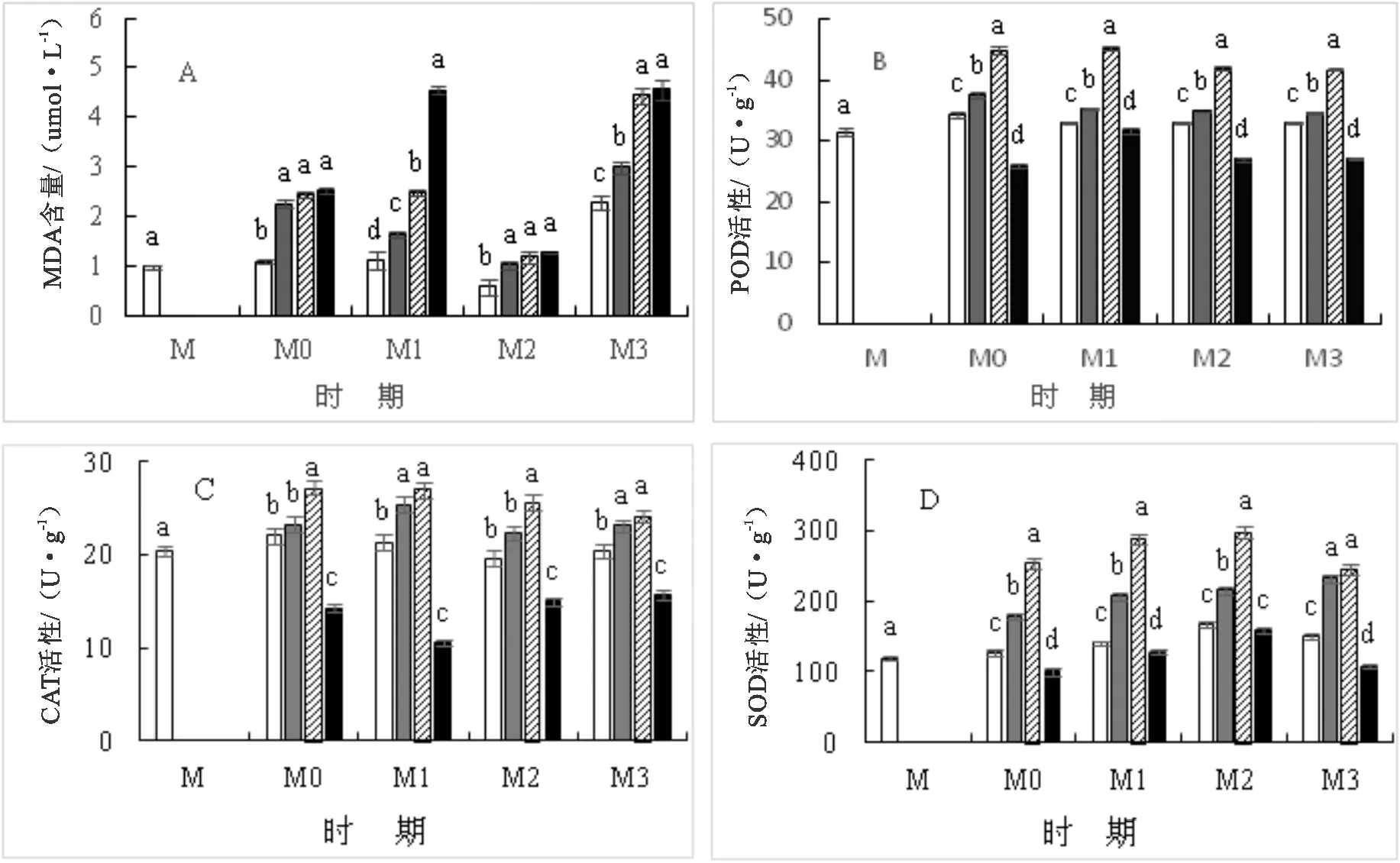
图3 不同浓度吲哚丁酸处理下的种子抗氧化酶活性
当IBA质量浓度为30 mg·L-1时,光皮桦种子体内的POD、CAT以及SOD酶活性最高,促进效果最显著(P<0.05)。在M0至M3的4个萌发时期中,30 mg·L-1的IBA处理光皮桦种子后,POD活性相比对照组分别增高23.5%、27.0%、21.2%、21.2%;CAT活性较对照处理组分别增高18.9%、21.0%、23.5%、15.7%;SOD活性较对照处理组分别增高49.9%、51.5%、43.9%、38.9%。
种子萌发中,随着IBA浓度的增加,种子体内的MDA含量增高(图3a);相同浓度IBA处理后,M3时期种子体内MDA含量高于其他时期;10、30、50 mg·L-1等3个浓度IAA处理下,同浓度M3时期种子的MDA含量与M0时期种子MDA含量相对比,分别增高25.0%、44.7%、44.7%。
3 讨论与结论
3.1 讨论
(1)外源激素处理与种子MDA含量的关系
种子萌发过程是种子与外界环境相适应、作用的过程,了解种子萌发过程中抗氧化酶活性变化有利于更深入和全面地获悉种子萌发特性[12]。植物器官在生长过程中受伤害时,会发生膜脂过氧化作用,最终分解产生丙二醛(MDA),因此MDA含量可作为评价细胞膜受损伤程度的重要指标[13]。
本试验中,光皮桦种子GA3处理后,萌发过程中体内MDA含量随GA3浓度增加呈不规则变化;IAA、IBA处理后的种子体内的MDA含量随GA3浓度增加呈增高趋势;低浓度(10 mg·L-1)GA3处理后的光皮桦种子,在露白时期(M1)体内MDA含量最高,种子细胞膜系统伤害最严重;在M0、M2、M3时期,随GA3处理浓度升高,种子体内MDA含量增高,对种子细胞膜的伤害加重,种子抗性逐渐降低。这与左月桃研究玉米种子GA3处理后胚内MDA含量呈现先增加后降低变化趋势结果[14]不同。
IAA、IBA处理后的光皮桦种子,体内MDA含量随着IAA、IBA浓度增高而增高,表明光皮桦种子细胞膜的损伤也随之加重。同一浓度IAA、IBA处理后的光皮桦种子在苗期种子(M3)时期MDA含量最高,说明在该时期植物细胞受到的损伤程度最为严重。以上说明,不同外源激素对不同种类种子萌发期间细胞膜产生的伤害程度不同,体内MDA含量的变化差异较大。
(2)外源激素处理与种子SOD、POD、CAT等抗氧化酶活性的关系
施加外源激素调控SOD、POD、CAT等抗氧化酶活性,维持种子体内活性氧的正常代谢和植物正常生长发育,可减缓发育过程中出现的膜脂过氧化对光皮桦种子细胞的伤害[15],可以使种子萌发后期生长力逐步增强,抗氧化酶活性增强[16]。
本试验表明,不同外源激素的不同浓度对光皮桦种子萌发过程中的抗氧化酶活性的影响存在差异,使用合适浓度外源激素处理种子,增强体内抗氧化酶活性使种子生长更迅速,种苗更健壮。GA3浸种处理后的光皮桦种子,萌发过程中POD、SOD、CAT酶活性随着处理浓度升高而增大,当浸种浓度达到50 mg·L-1时,种子体内的抗氧化酶活性最高,表明该浓度对抗氧化酶活性的促进效果最显著(P<0.05),与福建青冈种子GA3处理后子叶抗氧化酶活性变化规律一致[7]。
光皮桦种子在IAA、IBA浸种处理后的萌发过程中,SOD、POD、CAT酶活性随处理浓度增高呈先升高后降低的变化趋势。表明低浓度IAA、IBA对光皮桦种子体内抗氧化酶活性具有促进作用,高浓度IAA、IBA对种子体内抗氧化酶活性有抑制作用,与IBA溶液处理下山椒子幼苗叶片抗氧化酶活性变化趋势一致[17]。
3.2 结论
外部施用高浓度GA3和中低浓度IAA、IBA能显著提高光皮桦种子萌发过程中的抗氧化酶活性,促进种子生长,增强种子的生长力,提高幼苗的强健程度,有效解决光皮桦种苗培育和人工栽植上的一些困难,满足实际生产需求。
