EV71诱导has-miR-200b表达并促进自身复制的机制
2022-10-21胡静平傅煜轩
胡静平, 王 燕, 傅煜轩
(1. 苏州大学生物医学研究院, 苏州215123; 2. 南京医科大学附属苏州医院/苏州市立医院, 苏州215001)
肠道病毒71型(EV71)是一种非包膜、单链正义RNA病毒,属于微小RNA病毒科,是引发儿童手足口病(HFMD)的最主要病原体[1-2]。手足口病近年来在亚太地区广泛流行[3]。研究证实93%的手足口病死亡与EV71感染有关[4]。少数患者可发生无菌性脑膜炎、脑炎、急性弛缓性麻痹、神经源性肺水肿和心肌炎,进展迅速可导致死亡。目前并没有针对EV71病毒的特效抗病毒治疗方法,而EV71病毒的致病机制仍存在很多未知。
MicroRNAs(miRNAs)是一类较短的、高度保守的非编码RNA,由17~25个核苷酸组成,通过与mRNAs的3′非翻译区(3′-UTRs)的靶位点相互作用来抑制靶基因的表达[5]。大量证据表明miRNAs是病毒入侵和复制的关键调节器[6-8]。许多miRNAs已被报道参与病毒复制过程[9],例如,HIV-1衣壳结合宿主因子cPSF6在转录后受到细胞miR-125b的调控[10]。MicroRNA-135a通过下调宿主抗病毒因子调节丙型肝炎病毒基因组复制[11]。已有一些研究表明,宿主细胞中的miRNAs能够参与调控EV71的感染或复制。例如:EV71病毒通过调控宿主胞内miR-21的表达进而影响靶基因MyD88和IRAK1的表达,从而抑制宿主细胞的I型干扰素的产生[12];MiR-545通过靶向抑制PTEN和TRAF6蛋白表达从而促进EV71病毒复制[13];MicroRNA-302家族通过靶向KPNA2下调EV71诱导的先天性免疫应答[14]。这些发现都提示宿主细胞内的miRNA与EV71病毒的相互作用能够为抗病毒方面提供新的策略。为了寻找治疗EV71相关疾病有效的miRNA,还需要进一步研究miRNA与EV71感染之间相互作用的具体分子机制。
1 材料和方法
1.1 细胞和病毒
Hela细胞(人宫颈癌细胞)、SW480细胞(人结肠癌细胞)、293T细胞(人胚胎肾细胞)、Vero细胞(非洲绿猴肾细胞)均来自美国ATCC细胞库,在Dulbecco改良Eagle′s培养基(DMEM)(Gibco)中添加10%热灭活胎牛血清(Gibco),37 ℃,5% CO2。EV71(肠道病毒71型)由江苏省疾病预防控制中心馈赠。病毒溶液经Vero细胞培养扩增后,保存于-80 ℃冰箱中。病毒滴度由Reed-Münch终点计算方法确定[15]。
1.2 荧光定量PCR
使用Trizol试剂(Thermo Scientific)根据说明书进行细胞RNA提取。用primerscript RT试剂盒(Takara)将RNA逆转录为cDNA,根据样本数目按照说明书混合ABI SYBR Green Master Mix试剂,每管分别加入1 μL cDNA产物,在ABI 7500实时定量PCR仪中进行实时定量 PCR,反应程序设置:95 ℃预变性10 min;95 ℃变性15 s,60 ℃退火1 min,共计40个循环。每个基因做3个复孔,并重复3次实验,用DDCt法分析相对表达。miR-200b和U6引物购自RiBo生物技术有限公司(广州锐博生物技术有限公司),miR-200b表达以内源性U6 snRNA作为对照,IFN-β和EV71 vp1 RNA的表达以内源性GAPDH作为对照,目的基因相对表达量通过公式计算和分析,PCR引物序列见表1。

1.3 MicroRNA 转染
293T细胞接种于6孔板上,培养至细胞融合密度达到50%左右。根据说明书使用Lipofectamine 3000(Life Technologies,USA)将microRNA mimics或inhibitor(最终浓度为50 nmol/L)转染细胞。miR-200b mimics、mimic阴性对照(mimic-NC)、miR-200b inhibitor和inhibitor阴性对照(inhibitor-NC)均购自锐博生物技术有限公司。
1.4 蛋白质免疫印记
将裂解的细胞进行SDS-PAGE凝胶电泳,转移到PVDF膜上(Millipore,USA),并进行一抗、二抗孵育,结果通过Image J软件进行密度分析量化。使用的一抗为抗VP1抗体(Milipore,USA)、IFIT5抗体、GAPDH抗体(Proteintech,武汉)。
1.5 荧光素酶检测
从人类基因组DNA上PCR扩增IFIT5的3′UTR(核苷酸1212-1219),通过XhoI和XbaI位点(引物正向:5′-TCCTTCTTGGGTTCCT-3′;反向:5′- GAGGAGTCCAAATGCAAACTGG-3′)插入pmirGLO荧光素酶载体,构建野生型IFIT5 3′报告质粒,同时突变IFIT5 3′UTR通过PCR方法诱导点突变。293T细胞接种于96孔板中,转染含有IFIT5 3′-UTR和miR-200b mimics或mimic-NC的荧光素酶报告质粒。48 h后收集细胞,PBS洗涤一次,用裂解缓冲液(Promega)裂解。15 min后收集上清液,12 000 r/min离心5 min。用双荧光素酶报告检测试剂盒(Promega)测定荧光素酶活性。
1.6 免疫荧光
细胞感染后用3.7%多聚甲醛固定,用0.1% Triton X-100室温通透,依次用一抗和荧光二抗孵育,DAPI染料(碧云天,中国)用于细胞核染色。样品在激光共聚焦显微镜下观察并用成像系统自带软件进行分析(Olympus,日本)。
1.7 small RNA文库制备与miRNA测序
将感染及未感染EV71的细胞提取总RNA,每个样品取2 μg 总RNA进行small RNA文库构建。所有总RNA的RIN值都在8.0以上。根据Illumina TruSeq RNA Sample Preparation Kit(Illumia,USA)的操作说明分别选取不同的index标签建库。使用TruSeq PE Cluster Kit v3-cBot-HS(Illumia)试剂在cBot上生成簇。之后在Hiseq 4000测序平台上运行单端测序程序 (SE50),得到50 bp的单端测序reads。对各个样本中miRNA进行表达量进行统计,通过TPM进行表达量归一化处理,之后用DEGseq进行差异分析,将差异miRNA筛选条件设置为Pvalue<0.01,|log2(fold change)|>1。
1.8 统计学分析
2 结果与分析
2.1 EV71感染诱导宿主细胞内miR-200b表达上升
通过高通量测序检测microRNA在EV71感染SW480细胞中的表达情况。发现EV71感染SW480细胞24 h后,在所有统计差异变化的microRNA中,has-miR-200b变化最为显著[图1(a)和(b)];目前对miR-200b在EV71感染中的功能和作用未有报道。qRT-PCR进一步检测发现miR-200b的表达在EV71感染SW480细胞中不同时间点逐渐上升,18 h时达到峰值后不再变化[图1(c)];另外,不同MOI的病毒剂量感染SW480细胞后,发现与对照组细胞(MOI=0)相比,miR-200b的表达量随病毒剂量的提高而逐渐升高,当MOI=2时达到峰值后,不再随病毒剂量升高而增加[图1(d)]。
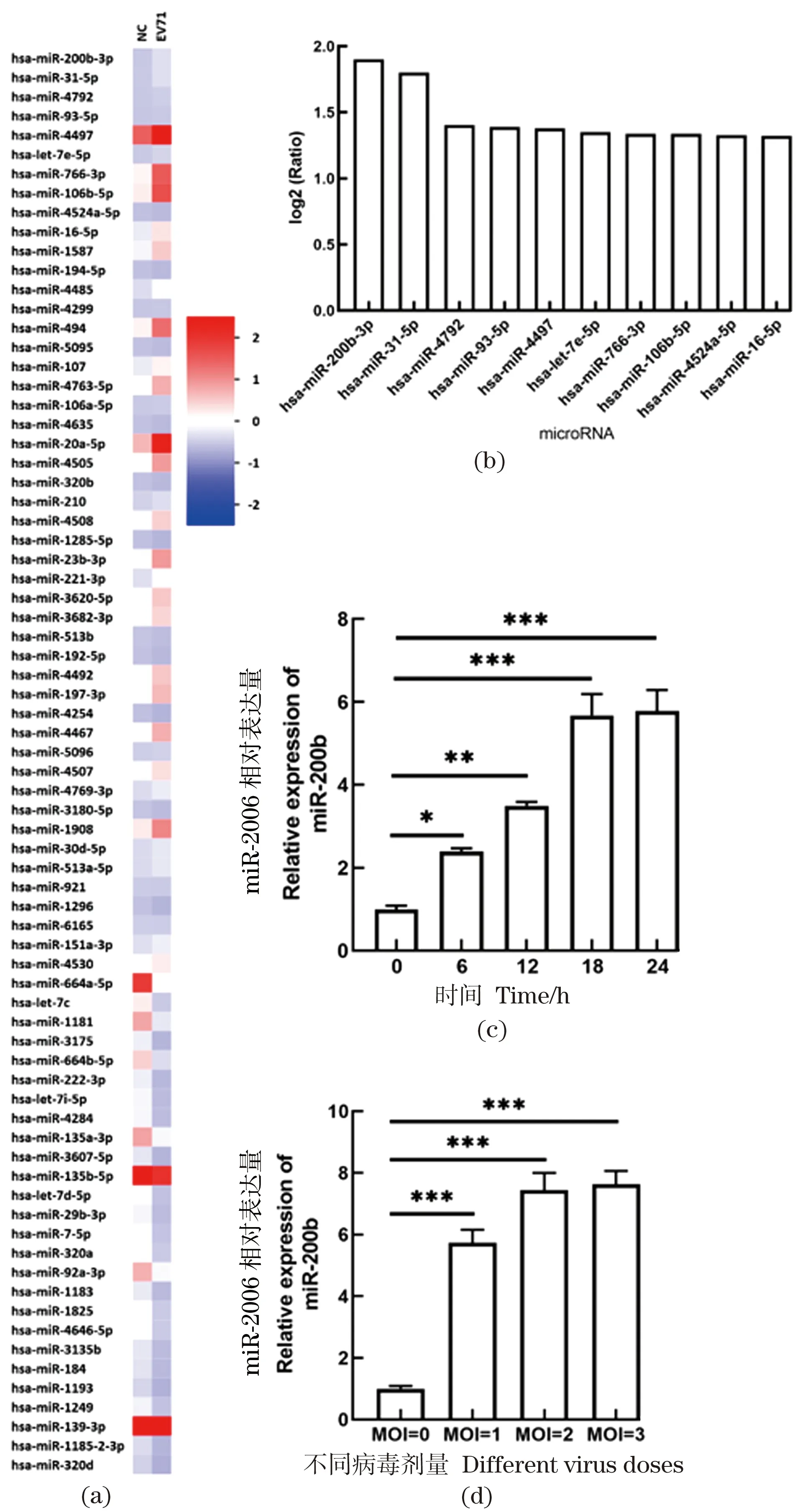
(a)在MOI=1条件下,EV71感染SW480细胞24 h时高通量测序分析结果;(b)高通量测序结果中升高倍数最高的top10 miRNA;(c)和(d)分别为用实时定量PCR定量分析EV71感染的SW480细胞不同时间点和不同病毒剂量后,细胞内miR-200b表达水平。* 表示P<0.05,** 表示P<0.01;*** 表示P<0.001。图1 在SW480细胞中,EV71感染上调miR-200b的表达Figure 1 EV71 infection in SW480 cells upregulates miR-200b expression
2.2 抑制miR-200b表达能够抑制EV71病毒复制
为了进一步验证miR-200b对EV71复制的影响,将miR-200b inhibitor和inhibitor-NC转染 SW480细胞24 h后,给予EV71感染24 h,检测病毒VP1的表达水平[图 2(a)],结果显示,与对照组或inhibitor-NC相比,miR-200b阻断可抑制EV71 VP1蛋白的表达。免疫荧光实验也证实了一致的结果,与阴性对照相比,抑制miR-200b的表达,HeLa细胞内EV71 VP1 蛋白表达显著降低[图 2(b)]。因此,阻断miR-200b可以抑制EV71的复制。
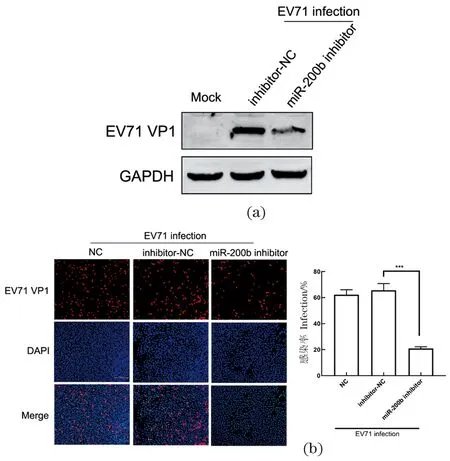
(a)转染miR-200b inhibitor或inhibitor-NC的SW480细胞感染EV71病毒后, Western Blot分析VP1表达水平;(b)免疫荧光法检测VP1蛋白表达。*** 表示P<0.001。图2 抑制miR-200b能抑制EV71复制Figure 2 Inhibition of miR-200b suppress EV71 replication
2.3 miR-200b能够靶结合IFIT5并抑制其表达
为探讨miR-200b调控EV71感染的分子机制,利用在线TargetScan软件进行生物信息学分析(www.targetscan.org),在预测的下游靶点中,IFIT5是miR-200b的潜在靶点之一[图 3(a)],IFIT5具有调节I型干扰素的功能,在抗病毒免疫中有重要作用[16]。为了证实IFIT5和miR-200b之间的相互作用,构建了含有野生型或突变型IFIT5 3′UTR序列的荧光素酶报告载体,并测定了其与miR-200b mimics或mimic-NC共转染细胞时的荧光素酶活性,如图 3(b)所示,与阴性对照相比,miR-200b mimics显著降低了293T细胞中含有野生型IFIT5 3′-UTR的报告基因的相对荧光素酶活性。相比之下,突变的IFIT5 3′UTR报告载体中的荧光素酶活性没有显示出显著的变化[图 3(b)],这表明miR-200b靶位点在IFIT5 3′UTR中为1212-1219。同时,过表达miR-200b会抑制IFIT5蛋白表达,而抑制内源性miR-200b,则会增加IFIT5蛋白表达[图 3(c)],表明miR-200b介导的IFIT5下调具有特异性。

(a)用TargetScan预测has-miR-200b结合位点(IFIT5 3′UTR及其突变位点);(b)荧光素酶报告基因检测,共转染含有has-miR-200b靶位点(wt-IFIT5 3′-UTR)或相应突变体(mt IFIT5 3′-UTR)的荧光素酶报告载体和miR-200b mimics或mimic-NC的293T细胞;(c)细胞转染miR-200b mimics、miR-200b inhibitor或阴性对照。24 h后,收集293T细胞,一抗:IFIT5和GAPDH抗体,EV71感染24 h,MOI=1。*** 表示P<0.001。图3 miR-200b能够靶结合IFIT5并抑制其表达Figure 3 miR-200b can target IFIT5 and inhibited its expression
2.4 miR-200b通过靶结合IFIT5进而抑制干扰素应答
IFIT5为重要的干扰素刺激基因,为进一步验证miR-200b是否能够通过靶向IFIT5来调节I型干扰素的表达,检测在IFN-β处理下,EV71感染SW480细胞后miR-200b的表达。荧光定量PCR分析显示,与阴性对照组相比,IFN-β的存在降低了miR-200b的水平[图 4(a)]。接着又检测了在miR-200b mimics或mimic-NC存在下,EV71感染SW480细胞中IFN-β的表达。荧光定量PCR分析显示miR-200b的过表达降低了IFN-β的表达水平[图 4(b)]。另外,通过抑制细胞内miR-200b表达后再给予EV71病毒感染,通过ELISA检测细胞上清液中IFN-β水平发现,在miR-200b被抑制的状态下,EV71病毒的感染会导致更多的IFN-β释放[图 4(c)]。这些结果表明miR-200b通过靶结合IFIT5进而抑制干扰素应答。
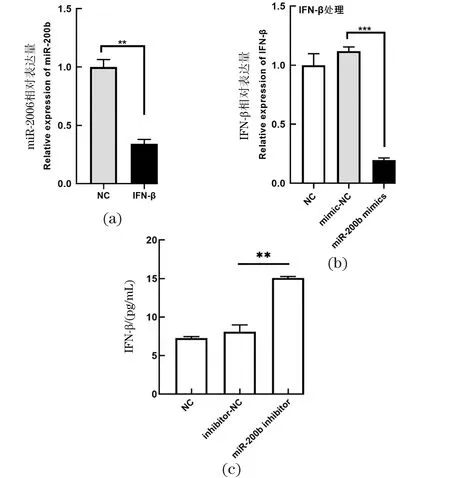
(a)在IFN-β(1 000 U)存在下,EV71感染SW480细胞,荧光定量PCR检测miR-200b mRNA水平;(b)EV71感染miR-200b mimics或mimic-NC转染的SW480细胞,实时定量PCR检测IFN-βmRNA水平;(c)ELISA检测在miR-200b表达被抑制下EV71病毒感染后的细胞上清液IFN-β的表达量。** 表示P<0.01;*** 表示P<0.001。图4 miR-200b通过靶向IFIT5抑制I型干扰素应答Figure 4 miR-200b inhibit the type I interferon response by targeting IFIT5
3 讨论与结论
EV71感染可引起一系列影响,从无症状感染到轻度手足口病,再到严重并发症,包括中枢神经系统和心肺衰竭,严重者死亡率高达82%~94%[17]。然而EV71感染的分子机制尚有很多未知,而且有效的治疗方法尚未开发出来。
MiRNAs在EV71病毒复制中起着重要作用,越来越多的证据表明,许多细胞MiRNAs参与了EV71感染和发病机制的调节,如miR-545[13]、miR-146a[18]、miR-494-3p[19]、miR-21[12]和miR-124[20]。本实验发现在EV71感染的SW480细胞中miR-200b表达水平升高。据报道,miR-200b是miR-200家族的重要成员。miR-200家族的成员已经被证明可以调节先天性免疫反应,研究表明miR-200b通过抑制c-Jun/MAPK通路降低活化小胶质细胞的炎症反应[21],为了进一步研究miR-200b在EV71复制中的作用,用miR-200b抑制剂下调HeLa细胞miR-200b水平,发现阻断miR-200b可以抑制EV71的复制。
为进一步研究机制,利用在线miRNA靶基因预测软件TargetScan对miR-200b的靶基因进行预测,发现IFIT5基因是miR-200b的靶点。IFITs是由干扰素诱导产生的含TPR结构域的蛋白,在机体抗病毒过程中起着关键作用[22-23]。IFIT5是一种干扰素调节因子,研究表明IFIT5在抗病毒反应中具有潜在作用[24]。我们发现miR-200b的过表达或内源性miRNA-200b的抑制分别导致IFIT5表达的抑制或增加,结果表明miR-200b介导的IFIT5下调具有特异性,进一步验证发现miR-200b的过表达会降低IFN-β的表达水平。
综上所述,EV71病毒感染能够上调miR-200b表达,进而抑制IFIT5基因表达,导致胞内干扰素应答抑制,从而有利于EV71病毒复制。结果为miR-200b在EV71病毒感染过程中的作用提供了一个新的机制,调控miR-200b的表达可能是应对EV71感染的一种潜在治疗策略。
