微量元素在林木中的应用研究进展
2022-10-19陈锐帆林嘉蓓邓小梅奚如春
张 恒,陈锐帆,林嘉蓓,邓小梅,奚如春
(华南农业大学林学与风景园林学院,广东省森林植物种质创新与利用重点实验室,广东 广州 510642)
植物体微量元素不仅参与酶、辅酶的构成,对叶绿素和蛋白质的合成及植物光合作用、代谢过程具有调节作用,还对大量元素的吸收和利用具有促进作用。如Zn能在300多种酶反应进程中起催化作用,并参与植物的呼吸和氧化过程、叶绿素和生长素的合成,以及碳水化合物的转化[1-2];Mn除了能调节光合作用,还在许多酶中作为辅助因子发挥结构作用[3];Fe参与植物体内呼吸作用、光合作用、DNA合成、氮素同化和固定、激素合成、活性氧的形成与消除等重要的生理代谢过程[4];Cu参与植物许多代谢过程,如线粒体呼吸、激素信号传导、光合电子传递,还是超氧化物歧化酶(SOD)的重要组分;Mo是另外一种参与植物代谢功能的重要元素,包括植物激素生物合成、嘌呤代谢、亚硫酸盐解毒和硝酸盐同化[5];B在植物中主要以B-糖复合物的形式存在于细胞壁中[6],参与半纤维素和细胞壁物质的合成,促进细胞伸长和细胞分化,促进生殖器官的建成和发育。因此,研究林木体内微量元素运输、分配和能量转化机制,并通过施肥手段为林木提供包含所需微量元素的养分,可以有效提高林木产量和改善林木品质[7-8]。
微量元素的研究已广泛应用于营养诊断以及林业施肥等各个方面,尤其是在农作物、蔬菜、果树等相关领域都取得一定的成果[9-12],但在林业领域的应用机制研究还有待加强。笔者基于近期有关林木微量元素的研究,从林木营养元素、微量元素与林木生长发育之间的关系及存在的问题等方面进行综述,并探讨其未来发展方向,以期为林木营养诊断及微量元素肥料的研究和利用提供科学依据。
1 林木施肥研究概况
德国在19世纪中叶最早重视林木对营养元素的需要和林地营养元素的循环研究,发现从林地收走枯枝落叶会导致森林生产力急剧下降,因此开始林木施肥试验[13]。但从世界范围来看,林木施肥一直处于试验阶段和小规模生产性施肥,直到20世纪50年代才开始在多个国家进行大面积推广。我国从20世纪70年代末开始林木施肥的试验,80年代以来研究了杉木(Cunninghamialanceolata)、桉树(Eucalyptusrobusta)、毛竹(Phyllostachysedulis)、青甘杨(Populusprzewalskii)、火炬松(Pinustaeda)和泡桐(Paulowniafortunei)等主要速生树种的施肥效应,90年代以来在杉木、桉树、欧美杨(Populus×canadensis)、毛竹、马尾松(Pinusmassoniana)、山核桃(Caryacathayensis)等用材林和经济林适生地区,进行了更加广泛的施肥效应试验,提出了针对不同树种的多种优化施肥方案,为生产上的合理施肥提供参考[14-15]。1991年,世界银行贷款国家造林项目“主要树种丰产林施肥技术研究与推广”和我国“八五”国家重点科技攻关项目“短周期工业用材林定向培育与维持地力措施研究”专题相继启动,此后10多年,我国把林木施肥列入国家科技攻关专题,针对纸浆材林微生物应用与施肥技术等专题开展了研究,这对推动我国林木施肥研究起到了积极的作用,也加快了林木营养元素研究的进程[16]。
2 林木微量元素分配及与林木各生长因子的关系
2.1 林木体内元素之间的关系
林木体内营养元素之间存在相互作用,即两种或两种以上元素之间能够产生促进或拮抗作用。国内外学者对元素之间的相互关联性做了大量研究,如在杨树体内,N、P同时存在会对Cu的含量有限制作用,P、B之间存在着拮抗作用[17],且P的含量过高时会引起微量元素(除Mn外)的紊乱和Cu、Zn的缺乏[18],而在缺Fe条件下,叶片Cu含量会显著上升。在缺P条件下,枳根系Cu含量上升而Fe含量降低,地上部分Zn含量上升而K浓度降低[19]。而对橄榄(Canariumalbum)的研究表明,缺K植株根系和地上部分Cu和Zn含量升高,K、Fe含量下降;缺Ca条件下植株根系和地上部分Fe、Zn含量增加,根系Cu含量降低[20]。因此,在研究营养元素与林木之间的关系时,需注意各种营养元素之间的相互关系。
2.2 微量元素与林木生长的关系
2.2.1 林木体内微量元素的分配
针对性地对林木进行营养研究,确立施肥最佳方案,必须首先了解微量元素的分布格局,明晰植株各器官对相关元素的需求。有研究发现,在橄榄胚胎发育阶段,除P和Na外,植株体内矿质元素Mg、K、Ca、N、Mn、Fe都处于最高水平,分化阶段的特征是K、N、Mg、Ga、Mn、Fe水平显著降低而Na元素显著增加,成熟期各元素水平均较低[21];水青冈(Faguslongipetiolata)植株中Fe、Zn、Al含量随根龄增长而增多,其中Zn在根茎、叶中的关联性最显著[22]。
营养元素在叶片中的含量大多高于其他部位的含量,说明叶片对元素的积累能力最强,且植物对这些微量元素的积累一般呈现Mn>Fe>Zn>B>Cu>Mo的顺序[23],如栾树(Koelreuteriapaniculata)人工林中,各器官微量元素含量大小顺序为叶>枝>细根>皮>粗根>大根>根头>干,各微量元素总含量顺序为Mn>Fe>Zn>Pb>Cu>Cd>Ni>Mo[24];油茶(Camelliaoleifera)林中,微量元素由高到低排序依次为叶、干、根、枝、花、果,其中Cu、Cd和Pb元素在枝中含量最高,Fe元素在根中含量最高[25]。Hagen-Thorn等[26]对几种温带树种的微量元素含量进行比较,发现叶中的含量大于根、茎(除Fe外),Cu的含量普遍最低,Mn在树种间差异最大且最易受土壤pH影响,另外对光合作用、蒸腾作用能动性影响大的元素如Fe、Mn在叶中会高于其他他部位;Kosiorek等[27]对欧洲赤松(Pinussylvestris)、白桦(Betulaplatyphylla)和挪威槭(Acerplatanoides)树叶和树皮中的微量元素含量进行测定发现,被检测树种的叶和树皮中其Fe、Mn和Zn含量明显高于其他元素。
2.2.2 微量元素对林木生长发育的影响
尽管植物对微量元素的需求量很少,但它们在植物代谢及生长发育中具有重要功能,缺乏任何一种微量元素都会影响植物的正常生长发育(表1)。如缺Zn植物会出现黄化和坏死斑点的形成,不利于林木生长发育[3, 28];缺Mn会引起缩果病并导致组织坏死和脉间黄化,阻碍植物生长[29-31];缺Fe会导致植物叶片脉间失绿降低林木生产力,如果不治疗甚至可能导致树木死亡[32-33];缺Cu主要影响生殖器官和幼叶,导致结实率降低,叶片顶端发白甚至顶梢枯死[34]。缺Mo症状是植株矮小,生长缓慢,叶片失绿且有大小不一的黄色和橙黄色斑点,严重时叶缘萎蔫或扭曲呈杯状,老叶变厚、焦枯,以致死亡;另外,缺Mo多发生在酸性土壤中,且伴有Mn和Al的毒害,常在酸性土壤上以施用石灰防止缺Mo症。B参与细胞分裂、细胞壁的合成及酚类化合物的变化等生理过程,此外B能促进花管萌发与其解毒作用密切相关,磷酸可与有毒物质黄碱素形成非活性物质使之失去毒性[3, 35];缺B会影响植物的营养生长和生殖生长,抑制细胞扩增,降低植物的繁殖力,并导致分生组织死亡[34]。Cl参与细胞质酶活动,帮助稳定膜电位,并在光合作用中发挥辅助因子的作用;缺Cl会造成叶子生长量减少和叶片枯萎坏死,严重时甚至会导致根系发育不良和果实变小[36-38]。Ni在许多酶结构起着关键作用,包括乙醛酶、尿素酶、超氧化物分解酶等;缺Ni会导致尿素积累、叶片坏死病变[39-41]。
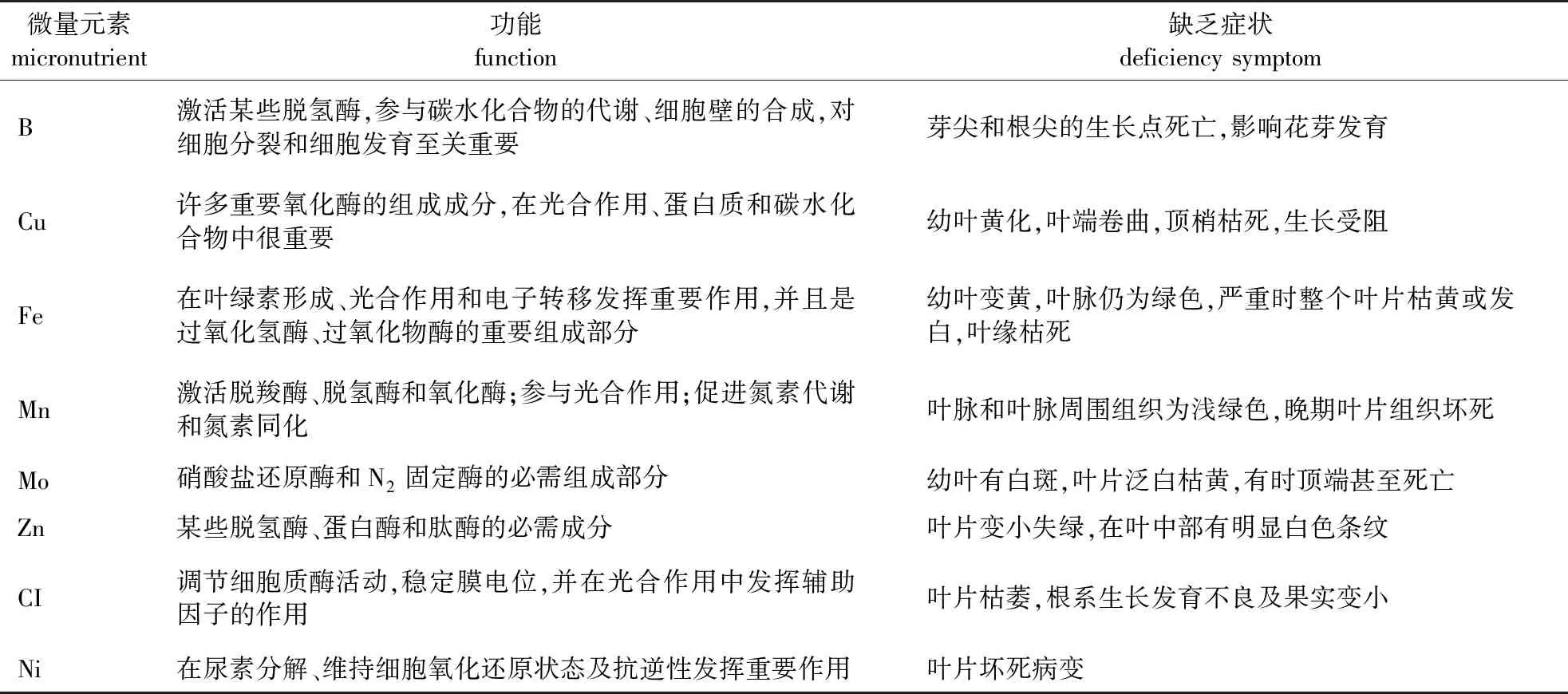
表1 微量元素在林木中的功能及其缺乏症状
已有研究表明,适当施用微量元素能促进林木生长发育,如:施用0.2%和0.3%的Zn对杜仲(Eucommiaulmoides)地径的增加均有显著促进作用[42];施用B肥能显著提高山核桃(Caryacathayensis)花粉活力,促进果实发育[43]。Göransson[44]研究在无污染的条件下Fe、Mn、Zn对垂枝桦(Betulapendula)的生长反应,结果显示,林木净增长与Fe、Mn、Zn的含量呈线性相关,在有Fe、Zn存在的条件下,林木对营养的需求是正常的多倍,但Fe、Zn超过一定含量时会维持在一定水平。
林木对不同微量元素及各元素间相互作用所产生的反应不同,如湿地松(Pinuselliottii)对微量元素反应的敏感度排序为B>Cu>Zn>Mn>Fe,在滨海平原对湿地松施用适量的Cu、Zn、Mn元素都会使芽质量增加,但微肥过量会造成Zn、Fe最先表现出毒害症状[17];Hoyle[45]研究了酸性土壤中生长的黄桦(Betulaalleghaniensis)对加入微量元素的反应,结果表明Cu、Zn、Mo的增加对植株的高度和胸径均无显著影响,但Zn对提高叶干质量有显著影响,Zn和Mn对提高茎干质量也有显著影响,而Mo、Cu、Zn对Al均有限制作用,且依次提高。林木与微量元素之间关系较为复杂,这可能与林木不同位置和不同生长发育阶段有关。Evans[46]测定了石梓(Gmelinachinensis)叶不同位置和不同叶龄的营养元素含量,结果发现随着树冠的增大,Zn、B含量降低而Fe含量增加,树冠内部的养分含量波动比外部轻微,叶龄的增加会导致Fe、B、Mn含量的增多。
2.2.3 林木对微量元素吸收转运的分子机制
微量元素参与非常广泛的代谢过程,包括从初级、次级代谢到细胞防御,从信号传导到基因调节、能量代谢和激素感知[47-49]。Fe、Zn是植物生长发育所必需的微量元素,在模式植物拟南芥(Arabidopsisthaliana)中,Zn调控转运蛋白家族(ZRT)及Fe调控转运蛋白家族(IRT)合称为锌铁转运蛋白(ZIP)。ZIP家族转运蛋白共有7个成员,分别为ZIP1~4和IRT1~3,其中ZIP1~3能在缺Zn的根中过表达,而ZIP4在缺Zn植物的根和芽中表达[50]。土壤中Fe3+由于其溶解度较低而无法在正常生理条件下被植物吸收。在非禾本科植物中,Fe的运输需要位于质膜的AHA分泌质子,从而增加根际酸化和促进Fe3+的溶解度,随后Fe3+通过还原酶基因FROs还原为Fe2+,被铁调节转运体IRT1基因转运到根细胞[51]。由于Fe2+转运蛋白IRT1对底物选择性不强,在转运Fe2+的同时也会非特异性地转运Zn2+和其他金属离子。NRAMP、ZIP和YSL转运蛋白家族可以促进植物对Mn的吸收,阳离子跨膜转运蛋白(CAX)、阳离子/钙交换剂(CCX)、P型ATPase、液泡铁离子转运蛋白(VIT)和阳离子扩散促进剂/金属耐受蛋白(CDF/MTP)可提高植物对锰抵抗的耐受能力[52-53],但目前对植物中Mn吸收的某些转运蛋白家族成员仍知之甚少。有研究发现,缺B条件下AdNIP5-1主要在猕猴桃(Actinidiachinensis)根部表达强烈上调,而在茎和叶中表达量较低,这对研究缺B植物生长发育及所需的B吸收至关重要[54]。枳缺B时,B转运蛋白PtrBOR3可在根中表达量升高,Ptr-BOR4在茎中表达量升高,而PtrBOR1、PtrBOR2表达量在根、叶中均升高[55]。关于Cu转运蛋白可分为3类,即吸收型Cu转运蛋白(COPT、ZIP和YSL蛋白家族)、排出型Cu转运蛋白(如HMA、PAA蛋白家族)和Cu伴侣蛋白(如CCS、ATX1和CCH)。其中对COPT蛋白家族的研究较多,植物转运蛋白COPT家族在橡胶树(Heveabrasiliensis)有9个成员,除HbCOPT2定位在线粒体膜内膜上,可能介导Cu2+由线粒体向胞质运输,其余均定位在细胞质膜上,推测它们可能负责吸收Cu2+,而大多HbCOPTs在橡胶树根部均有较高表达[56],所以COPT家族对调节林木体内Cu稳态平衡过程中发挥重要作用。Mo以各种形式存在土壤中,但只有钼酸盐(溶解形式)才能被植物吸收[57]。在模式植物拟南芥中,MOT1是位于质膜或内膜上的高亲和力钼酸盐转运蛋白,它可以帮助植物在土壤Mo含量很低的情况下吸收Mo[58-60];另一个拟南芥蛋白是液泡中的MOT2,在缺乏MOT2的植物中,其在叶片中的积累、种子中的减少以及衰老叶片中MOT2活性的增加表明,MOT2可能将钼酸盐从液泡中运输到细胞质中[61],这些研究表明钼酸盐转运蛋白能促进Mu的吸收和分配,从而保证植物正常生长发育。已被报道参与Cl-运输的转运体包括SLAC、ALMT、ABC、CLC、CCC和NRT等[62]。对柑橘的研究表明,NRT1基因在不含Cl-的基因中恒定表达,在Cl-积累基因型中表达下调,推测这可能与Cl-浓度过高对硝酸盐吸收的影响有关[63]。Ni可以通过各种阳离子运输系统运输,如Fe2+、Mg2+、Zn2+和Cu2+,也可以通过螯合剂、金属硫蛋白(MT)、YSL蛋白运输[64-66]。
2.3 微量元素与土壤、林分类型及林木病虫害的关系
2.3.1 与土壤的关系
植物微量元素含量与土壤养分密切相关,土壤中微量元素的有效性由母质、土壤的理化性质及生物因素的影响决定,如酸碱度、氧化还原电位、与共存离子的相互作用、与土壤矿物质的反应、有机质和土壤微生物活性等[67-69]。整地会改变土壤有效态微量元素(除有效B外)的剖面分布规律,使各土层中微量元素含量分布均匀[70-71]。丁锐等[72]研究发现各类土壤对B的吸附能力由大到小的顺序是砂岩类>花岗岩类>第四纪红壤类等>千枚岩类,对Zn的吸附能力由大到小的顺序是第四纪红壤类>千枚岩类>砂岩类>花岗岩类土壤,对Cu的吸附能力的差异较小。傅金和等[73]对杉木叶片微量元素含量与土壤的相关性研究表明,杉木叶片和土壤中均呈现Mn高量,Fe、Zn、Cu适量,B低量的现象,其原因是整个红壤区土壤有效B少(<0.5 μg/g)导致杉木叶中B含量低(<15 μg /g),在林地适当施用B肥后,发现对杉木根系发育及果实、种子形成会起促进作用,同时也证实了杉木是富集Mn的植物。土壤中微量元素的供给状况对植物的生长发育有着密切的关系,不同土壤中微量元素的含量、形态和分布规律有很大差别,对其进行合理分析和评价,有助于正确施肥。
土壤pH是影响林木对微量元素吸收的一个重要因子,因为它会影响土壤中有效微量元素含量。有研究发现,土壤有效B含量与土壤pH呈正相关,土壤有效Fe、有效Mn、有效Zn及有效Cu含量与土壤pH均呈负相关,而土壤中有效Mn含量与土壤pH相关性最大,有效Fe含量与土壤pH的相关性最小[74]。不同微量元素对土壤酸碱度的依赖性也不相同,如对蓝莓园的研究表明,Fe、Cu对pH的依赖性高于Zn、Mn,能被植物利用的有效Cu、Fe也高于Zn、Mn[75]。
土壤有机质与土壤微量元素有效态密切相关[76]。土壤有机质通过降低微量元素的活性,吸附、络合或沉淀土壤中的微量元素如Cu、Zn、Mn等,影响它们在土壤环境中的迁移转化[77]。曾曙才等[78]对宁镇丘陵区几种主要森林类型土壤中Fe、Mn、Cu、Zn、Mo、B等6种微量元素全量和有效量以及影响微量元素有效性的主要土壤因子等进行了研究,结果表明受土壤有机质和速效P含量等因子影响,该地区有效态B、Mo、Cu含量很低,均处于临界含量水平以下,有效Zn含量稍高,有效Fe、Mn特别丰富,Fe/Mn比例严重失调。含微量元素的土壤主要矿物质及pH对土壤和林木吸收微量元素的影响情况[79-83]汇总见表2。
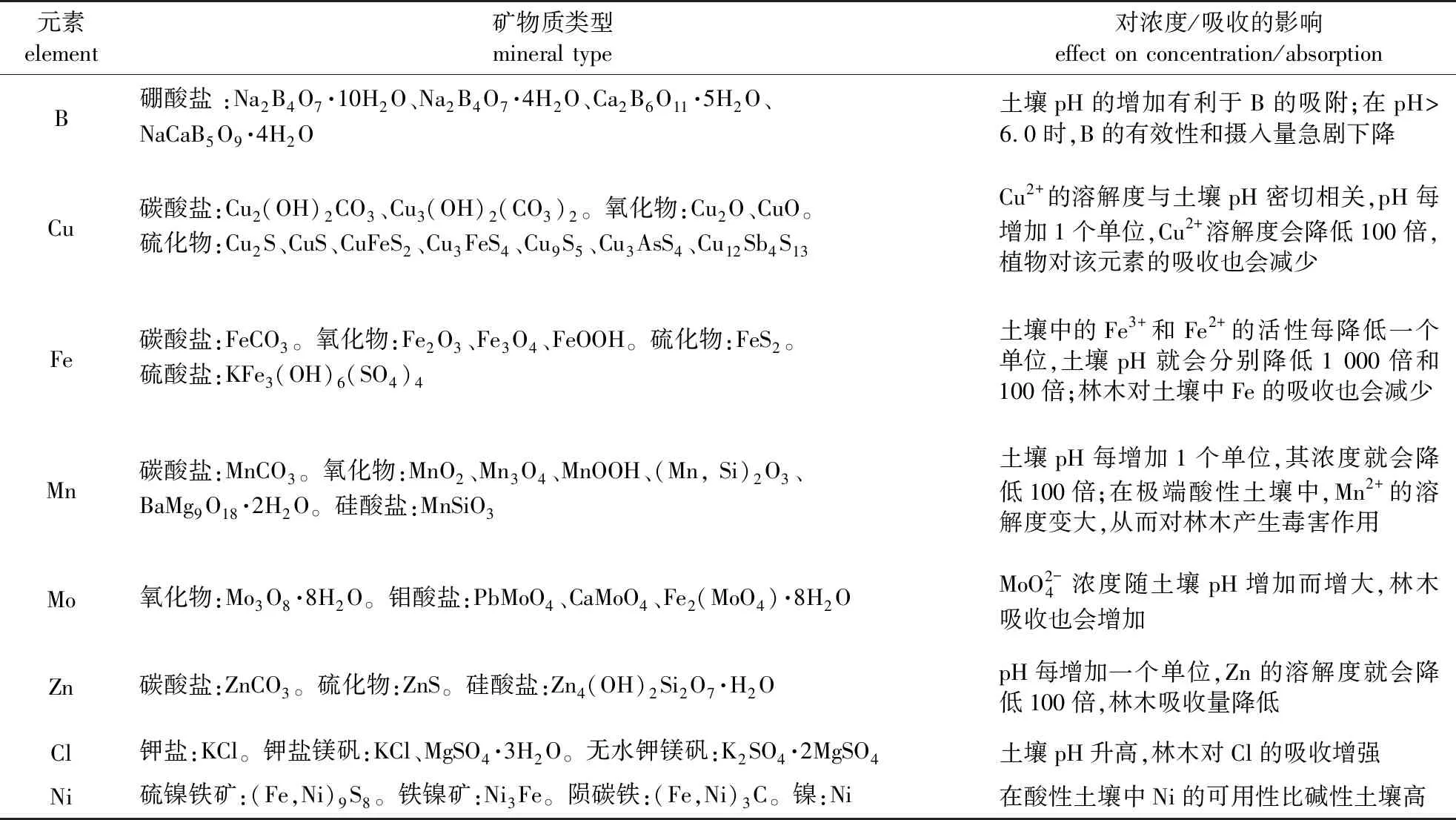
表2 土壤主要矿物质及pH对土壤和林木吸收微量元素的影响
2.3.2 与林分类型的关系
植物对营养元素的吸收和积累在不同林分中有一定区别。有研究表明,各混交树种营养元素含量存在较大差别,但营养元素含量比值变异系数较小[84],说明这些元素具有互补性,对树种生长具有协同作用,这种差别和互补性使种间关系向着协调方向发展,有利于林分生长。不同林分中,微量元素有效性综合指数大小为针阔混交林>灌木林>针叶林>阔叶林[85]。除混交外,轮作会影响林木元素水平,且Cu、Fe、Mn含量在轮作过程中含量变化最大[86]。混交和轮作能够提高林地营养元素的有效利用率,栽培2~3代桉树后接着栽培一代厚荚相思(Acaciacrassicarpa),能有效改良桉树林地土壤,恢复并提高土壤肥力[87]。林分密度也是制约微量元素有效含量的重要因素,如马尾松针叶凋落前,针叶中的Mn含量随林分密度增大而提高;凋落后,针叶中Fe转移率随林分密度增大而降低[88],这可能是因为针叶凋落前马尾松对Mn元素需求较多,而凋落后对Fe元素需求量较少。
通过了解林木在不同林分下的元素积累能力可以择优种植,提高效益。彭龙福[89]对不同立地条件下闽楠(Phoebebournei)人工林的乔木层、下木层、草本层、凋落物及分解物、土壤层进行取样检测,证实各营养元素的平均含量及其积累量均随林分立地质量下降而明显下降,营养元素在各层中积累量大小的总体趋势为乔木层>草本层>灌木层,所以改善林木类型可明显加快林地地力恢复和林木生长。
2.3.3 与其他环境条件的关系
林木生长的环境条件如病害、气候、物候期、化学物质等也常常会影响植株体内元素的含量,从而对林木生长发育产生影响。如对桑树(Morusalba)粗皮病与缺B情况的研究表明,干旱使B的有效性降低、固定作用增强,湿润多雨地区常由于强烈的淋洗作用而导致B的损失,且轻质土壤更加显著[90]。有时候砧木也会是树木缺素产生病症的因素之一,如苹果树(Maluspumila)嫁接用山丁子(Malusbaccata)做砧木易发生黄化,而以海棠(Malusspectabilis)、楸子(Malusprunifolia)做砧木则黄化相对较轻[91]。化学物质丹宁酸也会影响植株营养元素含量,丹宁酸含量较低时,营养元素释放多,且土壤表层5 cm处的含量比表层的要多,大量元素释放量比微量元素的高[92]。在板栗(Castaneamollissima)韧皮组织的营养变化和植物害虫的易感性研究中发现,与健康植株相比,染病植株中的N、K、Fe含量较高,Ca、N含量比值(记为Ca/N)较低,而健康植株中Ca/N不同时期有显著变化,Ca、Mg、K含量较高[93];对生长不良和健康生长的杧果(Mangiferaindica)树中营养元素的分布格局进行比较发现,在叶和嫩枝中,健康植株的Fe、Cu、Mn含量较低,生长畸形的植株叶和枝中Zn、B含量较高,花中微量元素含量普遍较低,新芽中较高[94]。
2.4 林木营养诊断及施肥方法的应用
2.4.1 功能性肥料研究与应用
为了提高肥料使用效率并最大限度地减少其对环境的不利影响,功能性肥料成为当前研究热点。功能性肥料是一种将营养物质与限制植物生长发育的因素相结合的具有针对性的新型肥料,主要是改善土壤理化性质、提高植物抗病虫害和抗逆性等能力的肥料。程文林等[95]对苹果喷施磁化复合肥发现叶片的Mn、Zn、Cu、Fe含量平均提高5.15%、10.80%、21.59%、20.51%;幼果的Mn、Zn、Cu、Fe含量平均提高27.18%、53.32%、19.73%、38.53%。王婷等[96]研究保水剂与外源物质复合配方对苗木蒸腾速率和叶水势的影响,筛选出山杏(Prunussibirica)、紫穗槐(Amorphafruticosa)的最佳配方分别为保水剂(SAP)10 g+吲哚乙酸(IAA)0.25%+萘乙酸(NAA)0.50%+聚天门冬氨酸(PASP)10 g和保水剂(SAP)15 g+IAA 0.25%+NAA 0.50%+PASP 1 g,这对干旱地区的苗木培育具有指导意义。广东省农业科学院从海洋生物、天然植物中提取了活性寡糖与多种微量元素复合而成的纯天然植物营养素应用于茶树(Camelliasinensis),结果表明,它对茶树生长有促进作用,增产可达7.9%~22.0%,且虫害减少,能促进茶叶品质的提高[97]。
2.4.2 微量元素施肥方法的应用
高效、节能地应用肥料已是一个重要的课题,多种功能性新型肥料在农林业发达国家已经广泛应用,发展中国家也在试图找出解决土壤缺素和作物生产力下降而又兼顾环境的施肥方法[98]。林木尤其是果树在正常生长过程中,对微量元素的需求量不多但极其重要。施用有机肥可使微量元素处于可利用状态并缓解其产生的毒性,大量元素的满足会促使微量元素的效应增加,因此要根据植物类型、各器官元素分布及土壤条件针对性地施用微量元素[99],还要根据土壤中微量元素含量、作物的营养特性、营养元素平衡及互作效应来选择合适的施肥方式[100]。
施肥时间和方法的合理结合也是提高肥效的重要途径,如在沙棘(Hippophaerhamnoides)园施肥要早施且少施氮,适时适量施用磷钾肥,多施有机堆肥,并注意施用中微量元素肥和微生物肥[101]。对柑橘的研究表明,喷施B肥后,叶的吸收量很少,仅0.3%,而且越老的部位营养积累量越少[102]。针对杧果缺B的习性,有研究指出在高降雨量地区,硼酸和农场厩肥结合施用效果理想,低降雨量区硼砂与农场厩肥混施效果佳[103]。微量元素既可通过土壤施肥,也可通过叶面施肥或种肥、底肥来调节,在颗粒状(或液态)大量元素中添加微量元素,可以使微量元素均匀分布,达到营养全面[104]。酸性土壤缺B可使用含B肥料如硼酸和四硼酸钠(Na2B4O7)来矫正;B肥可以与N肥和P肥混合使用,也可以单独施用,但要注意避免局部浓度过高产生毒害;浸种可用硼砂或硼酸溶液,根外追肥可用0.01%硼砂或硼酸溶液喷雾。对于多年生经济林,如苹果树、桑树等应推广叶面施B[90]。
3 林木施用微量元素存在的问题与展望
虽然我国存在林木可用的微肥品种和产量不多等问题,与国外相关情况相比还有一定差距,如尚未组织微肥生产和制定质量标准,产品未达到多样化、定型化等;同时,不同微量元素之间存在协同或拮抗作用,而目前生产中尚未掌握完整的土壤微量元素分布规律,林木施肥存在一定的盲目性。随着我国生态农林业的发展,微生物肥料是目前研究热点,它具有无污染、更高效的优点。因此在林木施肥中采用微生物肥,可以减缓土地营养衰退趋势,同时通过树种选优、改良土壤、加强管理来提高总体效益。
1)考虑客观条件进行试验。根据以上的总结,有关微量元素的实验很多是在无病毒、不受干扰或标准立地条件下完成的,其结果和实际情况有一定的差距,因此生产中需要大田试验才可推广。另外,由于物候期的不同,试验时要提前多次观察标记并保证植株健康,且各试验目标处在同一水平上。处于幼龄期的林分,由于没有和生长状况紧密结合,其中很多因子还未呈现出规律性,再加上土壤营养元素含量积累量、林下植被、理化性质的变化需要进一步研究。因此,肥料的实施和分配,在实际环境下应根据不同条件不同对待。
2)充分运用地被物的养分循环。很多研究过程都涉及凋落物的营养元素诊断,作为营养元素生物循环的主要归还者,凋落物可以促成土壤腐殖质的合成和分解,加快和补充林地养分的循环和积累,改善土壤物理性质,提高林分水源涵养功能。如杉木林净生产力(5.298 t/hm2)是活地被物(1.356 t/hm2)的4倍多,但它们的微量元素年净积累量相差无几,杉木为2.274 kg/(hm2·a)、活地被物为2.145 kg/(hm2·a),由此看来,活地被物对营养元素的吸收能力较强,故可以每年在成林里对活地被物进行刈割,置于林地以加速林木的养分循环,并可缓解活地被物与林木竞争养分[73]。
3)定向培育。根据微量元素在林木不同器官的含量和各元素之间的相互作用,可以针对性地实施对所需部位生长有利的施肥方法提高林木经济效益。我国在人工林经营上笼统地强调“速生丰产”,没有具体的生产目标,盲目造林和盲目施肥会造成人力、物力、财力上的极大浪费。施肥在增加林木生长、提高木材产量的同时,也会影响林木抗霜冻能力、抗病虫能力,以及木材的材质、材性和木浆质量等,因此作为不同培育目标的同一树种施肥技术应是不相同的。另外,植物叶片缺素临界值和土壤有效元素临界值的测定十分必要,植物的缺素症状并非都能表现出来,有些未表现的潜在缺素症状可能占据更大的比例[105],要针对性地观察和试验才能确定林木对元素的具体需要。施肥作为林木培育的一种基础技术措施,应该在不同的培育目标下进行分类研究。
4)诊断技术的系统化与程序化。林木营养诊断标准是搞好林分营养管理的一个有效工具,除受养分元素本身的影响外,还受环境因子和林木生长,以及人为因素如研究区域、采样数量、分析方法和研究手段等的影响。林木种类不同及生长发育阶段不同对营养元素的需求也不尽相同,我国现有的一些营养诊断标准只是一种局部的结果,因此可对这方面养分需求规律进行研究。另外,可将营养诊断技术与计算机技术融合来指导林木的合理施肥。
5)利用生物技术提高林木对微量元素的利用率。虽然目前一些关于肥料的研究成果已经应用于实践,但是就国内来说各方面还比较欠缺。如对微量元素研究的分子机制多为模式植物拟南芥,针对林木的研究较少。因此运用先进的生物技术更加深入地了解树木养分代谢机理、营养吸收特性及需肥规律极为重要,如通过分子生物学探究林木体内对微量元素吸收或耐受的基因,还可以利用微生物学,如菌根真菌、细菌等提高林木在逆境下(微量元素浓度过高或过低)的适应性并在实际中应用。
