黄栌光合和呈色特性对重庆阴雨天气的响应
2022-10-18李林珂王一诺吴焦焦荣星宇段儒蓉
李林珂,王一诺,薛 潇,张 文,吴焦焦,高 岚,谭 星,荣星宇,段儒蓉,刘 芸*
(1. 西南大学资源环境学院,重庆 400715;2. 重庆市黔江中学,重庆 409000)
黄栌属(Cotinus)树种分布于我国华北地区和长江流域,是我国秋冬季重要的风景林彩叶树种[1-4];其根系发达、萌蘖力强,也是干瘠山区造林绿化的优良先锋树种[5-8]。三峡红叶景观是长江三峡重要的旅游资源,主要由漆树科(Anacardiaceae)黄栌属树种叶色变化形成。黄栌(Cotinuscoggygria)在三峡库区的生长节律为:3月萌芽、展叶,4—5月开花结实,6—8月进入营养生长旺盛期,10月末叶片进入变色期,至12月末变色结束进入落叶期[9]。由于该属树种分布面积大(长江三峡两岸黄栌集中分布面积约2 万hm2)、观赏周期长(40余天),在库区农民脱贫致富中发挥了积极的社会效益和良好的经济效益。
气候变化对植物生长发育的影响随植物种类、环境条件而异[10-13]。黄栌是阳性植物,喜光,耐干旱贫瘠[5-8,14-21],光照充足环境的黄栌叶片鲜艳美丽[3-4],干旱胁迫环境下黄栌幼苗叶片更薄、更小[20-21]。目前关于气候对黄栌影响的研究主要集中在光照方面[2-5,14-15,17-22],而关于温度、湿度等其他气候因子对黄栌影响的研究较少,且研究结果存在分歧。如葛雨萱等[23]发现湿度对黄栌叶片变色影响不大,但李海龙[24]认为适当增加湿度可缓解黄栌叶片花青素减少,提高观赏价值,而高温条件下不利于黄栌叶片色彩的表达。重庆市气候类型为亚热带湿润季风气候,夏秋两季(立夏至冬至)的降水量占全年总降水量70%左右,年平均相对湿度为70%~80%,属于中国高湿区;年日照时间1 000~1 400 h,日照天数占比仅25%~35%,是中国全年日照最少的地区之一[26]。为全面研究气候因子对三峡库区黄栌生长发育的影响,以黄栌为材料,探讨天气变化对黄栌光合作用和叶片呈色特性的影响,以期为苗木引种和科学抚育提供依据。
1 材料与方法
1.1 研究区概况及供试材料
试验地位于重庆市西南大学竹园后山(106°25′54″E,29°47′56″N),海拔227 m,相对高度50~100 m,坡向正南,立地类型为低丘,该区域属于典型的亚热带季风气候,2019年4—11月重庆天气阴雨连绵,日照时间共80.58 h,平均相对湿度75.81%。2016—2019年有关气象数据购自重庆市北碚区气象局,用于气象数据时空分布差异分析。
以4年生黄栌实生苗为材料,采用露天盆栽方式(栽培土壤选自试验园),选取长势一致、无病虫害的黄栌48株,种于规格为30 cm × 40 cm(直径×高)控根容器内,分为6组,每组8株,挂牌标记,实验期间不施肥、不浇水。2019年4—11月于每月中旬选择连续晴天时进行样品采集和指标测定。每次采样时,从每组材料中随机抽出未曾取样的植株1株,共6株,在同一方位选取枝条顶端往下数第3~5片健康成熟叶进行光合指标测定并采集叶片。
1.2 指标测定方法
1)光合指标测定。采用Li-6800便携式光合作用测量仪(LI-COR,USA)测定叶片净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)和气孔导度(Gs)。
2)光响应曲线拟合。采用Li-6800便携式光合作用测量仪红蓝光源,使用CO2钢瓶作为CO2来源,控制CO2含量为400 μmol/mol,温度(T)设置25℃,相对湿度(RH)设置65%,选择完整、干净的待测叶片并做好标记,测定不同光照强度下黄栌叶片的Pn。测定时间为09:00—11:00,测定前以1 800 μmol/(m2·s)的光强诱导待测叶,并设置10个光强梯度为1 800、1 500、1 200、900、600、300、200、100、50和0 μmol/m2,依次进行测定,重复3次。使用“光合计算4.1”软件[27]进行光合-光响应曲线拟合。
3)CO2响应曲线拟合。待测叶片选取位置与光响应曲线测定时的相同,将测定光强设置为近乎叶片饱和光强时的光照强度,其他控制条件和测定光响应曲线保持一致,使用CO2钢瓶作为CO2来源,测定时仪器自动匹配浓度,CO2含量梯度设定为1 800、1 500、1 200、900、600、300、200、100、50、0 μmol/mol,重复3次。CO2响应曲线采用双曲线修正模型[27]进行拟合。
4)叶绿素含量的测定。将采集的叶片装入内有冰袋的保鲜袋及时带回实验室,采用乙醇浸提法[28]用可见分光光度计(Spectrumlab 22)测定并计算叶绿素总量(Chls)和类胡萝卜素(Car)含量(mg/g)。
5)花青素(Ant)含量的测定。采集叶片后,参照文献[29]方法,用打孔器取新鲜叶片0.03 g,每个处理重复3次,将称取的叶片放入25 mL容量瓶中,加入10 mL的1%盐酸-甲醇溶液,将容量瓶封口,充分摇匀使叶片完全浸入提取液中,放于避光处浸泡24 h至叶片无色,得到花青苷提取液。所得浸提液用紫外分光光度计在波长600 nm和530 nm下比色,以“U”表示一个花青苷单位,则以每克鲜叶提取液的光密度变化值λOD530 nm-λOD600 nm=0.1 U。
6)叶色参数测定。采集后的黄栌叶片平整固定于纯白色背景纸,使用Canon EOS-7D数码照相机进行拍照并确定叶片颜色参数L*、a*、b*值[30-31]。每片叶均在避开叶脉处的叶尖部、叶中部、叶基部相同位置各取2个色点,共6个色点,将6个点的平均值作为该叶片的叶色参数。
1.3 数据处理
用Excel 2010进行数据统计与图表制作,用SPSS 25.0进行差异性分析与相关性分析,以及对光响应曲线、CO2响应曲线的拟合。采用t检验法比较数据组间差异(α=0.05),数据用平均值±标准误差表示。
2 结果与分析
2.1 2016—2019年试验地气候年际动态
重庆北碚区2016—2019年黄栌生长季(4—11月)逐月月平均气候数据统计见图1。从图1可以看出,2019年4、7、10月的月平均降水量(AMP)比2016—2018年的高出16%~64%;2019年5、7、10月日降水量≥0.1 mm的日数(DPD)比2016—2018年多出3%~11%;2019年5—7月的月平均相对湿度(RH)比2016—2018年的高出4%~16%;2019年5—7月的月平均日照时间(SD)比2016—2018年的少24%~60%,7月的月平均日照时间比2017—2018年的少53%~57%;2019年5、7月的月平均气温(Q)比2016—2018年的低6%~11%。
重庆北碚区2016—2019年的4—11月气候数据统计见图1。与2016—2018年的4—11月相比,2019年4—11月日照时间减少10%~25%,月平均降水量高出12.5%~25.0%。2019年4—11月的降雨总天数超过55%,与该区域常有的天气状况比较,5—8月湿度大、降雨多、光照强度弱;与2016—2018年相比,2019年阴雨天气较多。
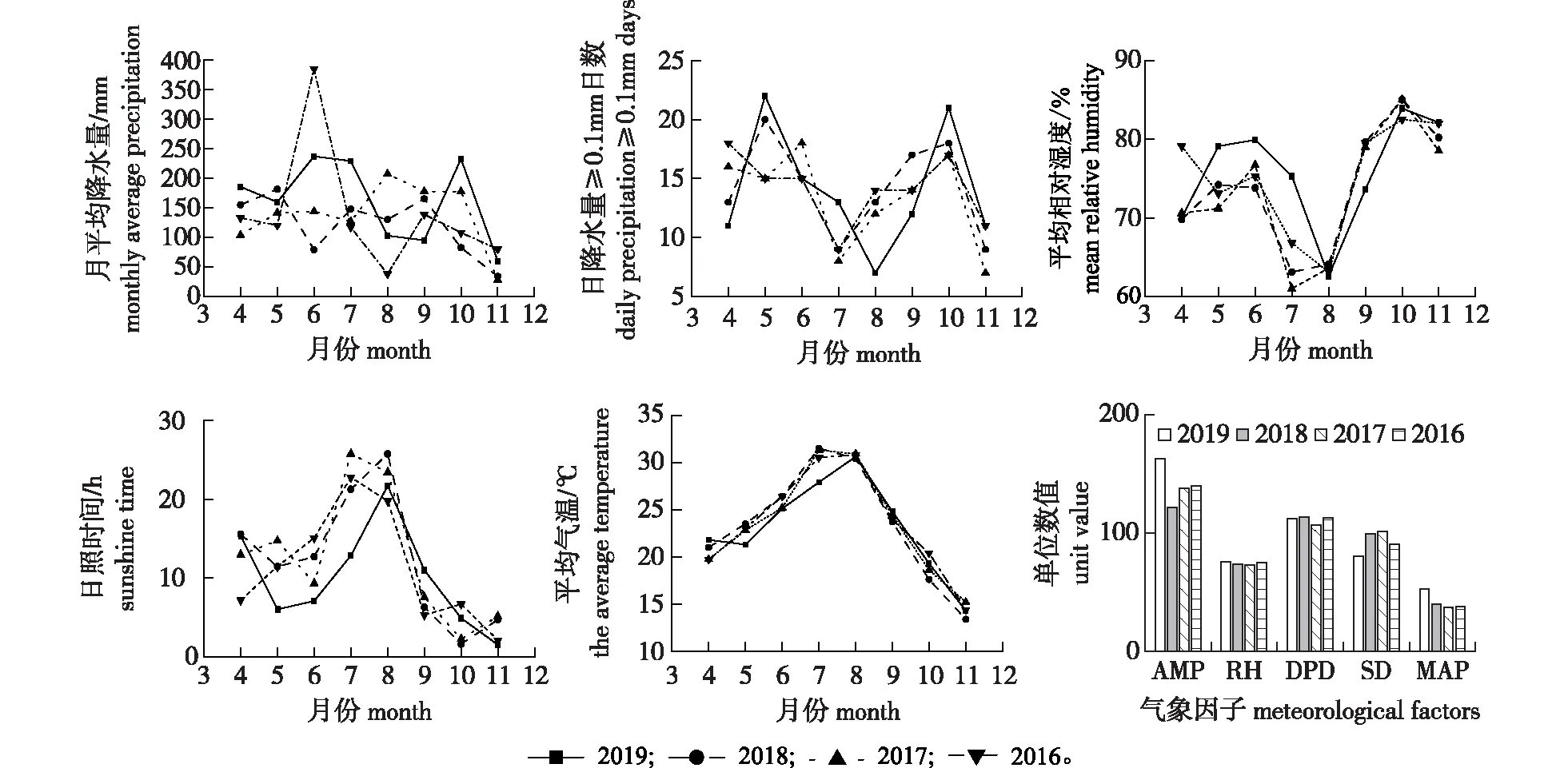
AMP.月平均降水量monthly average pecipitation;RH.相对湿度relative humidity;DPD.日降水量≥0.1 mm日数daily precipitation ≥0.1 mm days; SD.日照时间sunshine duration;MAP.平均最大日降水量arerage max daily precipitation。下同。The same below.图1 重庆北碚区2016—2019年4—11月逐月气候数据及气象因子统计Fig.1 Monthly climate data statistics and meteorological factors of Beibei District in Chongqing from April to November,2016-2019
2.2 阴雨天气对黄栌叶片色素含量及光合参数的影响
2.2.1 2019年4—11月叶片色素含量与光合参数动态
2019年4—11月黄栌叶片色素含量动态见表1。从表1可知,叶绿素a(Chla)含量从4月开始逐月下降,至9月显著上升(P<0.05),后又呈下降趋势。叶绿素b(Chlb)含量从4—6月显著下降,7—9月缓慢上升。类胡萝卜素(Car)含量在9月最高,5月次之。花青素(Ant)含量变化趋势与Chla相似。在重庆地区,5—8月为黄栌生长旺盛期[9],在此期间,成熟叶片中叶绿素(Chls)含量达到最高值。分析认为,2019年5月以来的连绵阴雨天气,使得叶片色素合成受阻, 8月天气转好,光照充足(图1),有利于色素合成,9月所测4种色素含量均与天气条件变化有关。观测期内,黄栌光合参数变化符合一般规律,但也表现出受天气条件的影响较大(表1)。

表1 2019年4—11月黄栌叶片色素含量与光合参数动态
2.2.2 黄栌色素含量、光合参数与气候因子的相关性分析
黄栌色素含量、光合参数与气候因子的相关关系见表2。由表2可知,Chla含量与平均气温(Q)显著正相关(P<0.05),与其他气候因子相关性不显著(P>0.05);Chlb含量与各气象因子相关性不显著;类胡萝卜素(Car)含量与月均降水量(AMP)极显著负相关(P<0.01),与其他气候因子相关性不显著(P>0.05);花青素(Ant)含量与月平均降水量显著负相关(P<0.5)。
Pn与平均气温(Q)、日照时间(SD)极显著正相关(P<0.01),与光合有效辐射(PAR)显著正相关(P<0.05),与日降水量≥0.1 mm日数(DPD)极显著负相关(P<0.01);Tr与SD、Q极显著正相关(P<0.01),与PAR显著正相关(P<0.05),与DPD极显著负相关(P<0.01),与大气二氧化碳浓度(Ca)显著负相关(P<0.05)。

表2 黄栌4—11月色素含量、光合参数与气象因子的相关性

图2 黄栌的Pn-PAR与Pn-CO2响应拟合曲线Fig.2 The response fitting curve of Pn-PAR and Pn-CO2 of Cotinus coggygria
2.3 黄栌4—9月的Pn-PAR与Pn-Ci响应过程
通过Thornley模型[32]拟合,得到黄栌4—9月的Pn-PAR曲线拟合效果图(图2A),同时计算得到黄栌最大净光合速率(Pn,max)为(5.346±0.682)μmol/(m2·s),暗呼吸速率(Rd)为(0.532±0.061) μmol/(m2·s),光补偿点(LCP)为(30.654±6.642) μmol/(m2·s),光饱和点(LSP)为(1 511.211±122.642) μmol/(m2·s),表观量子效率(AQY)为(0.019±0.002) mol/mol。以黄栌生长旺盛期(8月)所测数据为例,黄栌光合作用的光响应过程为:当光照强度在0~300 μmol /(m2·s)时,Pn随着PAR的增强逐渐上升,当PAR达到1 511.21 μmol /(m2·s)时,Pn达到最大值,之后随着PAR继续增强,Pn趋于平缓。LCP能够反映植物对弱光的适应能力,LSP能够反映植物对强光的适应能力[33],最大净光合速率(Pn,max)反映叶片光合能力的强弱[34]。阳、阴性植物主要反映植物对光照强度需求的差异。阳性植物在强光环境中生长发育健壮,在弱光条件下生长发育不良,有较高的光补偿点[40.5~81.0 μmol/(m2·s)]及光饱和点[1 620~2 025 μmol/(m2·s)];阴性植物在弱光下比强光下生长好,有较低的光补偿点[约8.1 μmol/(m2·s)]及光饱和点[405~810 μmol/(m2·s)][35-36]。与大多数阳性植物相比,黄栌具有较宽的光能利用幅度,其LSP 为1 511 μmol/(m2·s)、LCP为30 μmol/(m2·s),表明其在强光、弱光环境下均具有较强的耐受能力。在光照充足、降雨较少的8月,黄栌AQY明显高于其他各月,而在出现连绵阴雨天气的4—7月,黄栌AQY显著下降,尤其是降雨最多的6月。9月后黄栌叶片逐渐进入衰老期,LCP轻微下降,LSP明显升高,光响应随连绵阴雨天气的变化说明黄栌从正常生长变为耐受适应。
根据双曲线修正模型的拟合结果,得到饱和CO2环境下的同化速率(Amax)为(4.287±0.043) μmol/(m2·s),而CO2补偿点为(29.383±9.378) μmol/mol,CO2饱和点为(1 548.030±4.029) μmol/mol。黄栌光合作用的CO2响应过程见图2B,以8月为例,当CO2含量在0~300 μmol /mol时,Pn曲线几乎为线性增长,随CO2浓度的增大逐步上升,达到饱和点后,Pn趋于稳定。Pn与Ci之间呈正相关,这种正相关说明,光合速率的增高是Ci增高的结果,是两者关系的规律性反映[37]。研究表明,C3植物的CO2饱和点一般在1 000~1 500 μmol/mol,CO2补偿点一般在40~70 μmol/mol[38],而黄栌的CO2饱和点为1 540 μmol/mol,CO2补偿点为29 μmol/mol,高的CO2饱和点反映黄栌具有强的光合能力,低的CO2补偿点反映出黄栌较强的碳同化能力[39-40],可见,在连绵阴雨天气下黄栌较宽的CO2利用幅度有利于体内有机物的积累。CO2响应曲线变化过程与光响应曲线变化相似,遭遇连绵阴雨天气后,曲线变化随连绵阴雨天气的变化呈现为CO2饱和点降低,光合能力降低,CO2补偿点变化不大,Amax随阴雨天气的增多而下降的趋势。
2.4 黄栌叶色参数动态与分析
在Lab模型中,L*代表亮度,L*值越大表示颜色越明亮;色相参数a*表示红/绿属性色相,色相参数b*表示黄/蓝属性色相,即a*(b*)值越高,红色(蓝色)越深[31,41]。由表3可见,黄栌叶色L*值在8—11月表现为降低;a*值在5—10月呈上升趋势,11月略有下降;b*值在8—10月上升,11月下降。色相角[h=argtan(b*/a*)]在8—10月增大,11月下降,且在各时期差异均不显著(P>0.05);叶片彩度(C*)值自8月至10月增大,11月下降,其中11月与9月、10月差异不显著(P>0.05)。在重庆地区,黄栌叶片转色期从10月份开始,11月为转色峰期,至11月末颜色最为亮丽[26,31]。本研究中黄栌叶色在10月份最亮,11月份则呈现暗红甚至褐色。
黄栌叶色参数与色素含量、光合参数之间的相关关系见表4。由表4可知,L*与Chlb含量极显著正相关(P<0.01),与Chla含量显著正相关(P<0.05);a*与Chla含量、Chlb含量极显著负相关(P<0.01);其他色素含量与叶色参数间相关性均不显著(P>0.05)。a*与Tr、Pn、Gs极显著负相关(P<0.01);b*与Tr、Pn、Gs极显著负相关(P<0.01);C*与Tr、Pn极显著负相关(P<0.01),与Gs显著负相关(P<0.05)。
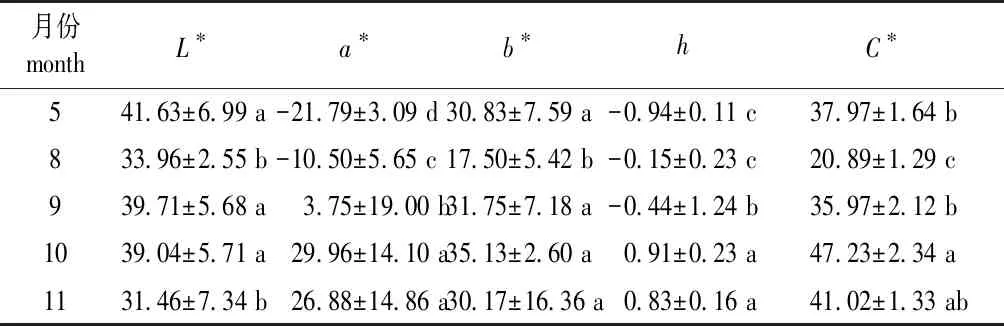
表3 不同时间黄栌叶色参数

表4 叶色参数与色素含量、光合参数之间的相关性
3 讨 论
光合色素含量高低在一定程度上反映植物同化物质的能力[42]。杨逢建等[43]发现光照强弱对植物光合色素形成、含量和分布产生直接影响,阴天不仅光照弱,而且蓝光和绿光成分增多[44],Chlb可以吸收Chla不能吸收的470~650 nm波长处的光能。陶俊等[45]发现,长期弱光下生长的植物,不仅Chls降低,类胡萝卜素含量也随之降低。本研究同样表明,连绵阴雨天气导致阳性植物黄栌光合色素合成减少,但Chlb含量却相对增加,尤其在Chla降幅较大的6—8月,Chlb呈小幅度增加。吴焦焦等[9]对同一批黄栌的研究结果也证明,在光照较好的天气状况下,Chls、类胡萝卜素含量、花青素含量更高。郭春爱等[46]认为植物能根据不同的环境条件,利用叶绿素循环来调整Chla与Chlb的比值,当植物生长在弱光下时,Chlb的合成加快,捕获更多的光子转移到Chla,这是植物生长对弱光环境的适应机制[47]。有研究表明,弱光还可以抑制或下调基因的表达,抑制花青素苷的合成[48]。而在光照充足的情况下,较高的Chls含量有利于提高叶片的光合效率,促进可溶性糖积累,为花青苷合成提供充足原料,黄栌叶片呈现亮丽的色彩[3-4, 9,27]。黄栌呈现红色是由于其花青素苷(主要是矢车菊素3-O-半乳糖苷)的相对含量增加,而叶片艳丽光亮则是因为黄酮醇苷的积累[3-4,31]。葛雨萱等[3-4]在控制实验下发现黄栌在低光照环境生长受限,植株矮小,叶片稀疏,呈色效果差,位于山顶光照充足的叶片鲜艳美丽。本研究发现,花青素含量与日降水量≥0.1 mm日数(DPD)显著负相关(P<0.05),而L*与Chlb含量极显著正相关(P<0.01),与Chla含量显著正相关(P<0.05),a*与Chla含量、Chlb含量极显著负相关(P<0.01),说明随降水日数(阴雨天气)增加,光照时间相应减少,导致黄栌生长光照不足,影响其Chls合成,降低Pn[25],有机物合成减少,不利于花青素苷和黄酮醇苷积累[3-4,29],最终影响叶片呈色质量[24]。可见,重庆地区2019年遭遇的连绵阴雨天气是导致黄栌叶片呈色质量较差的主要原因。
光照和降雨量是连绵阴雨条件下影响黄栌光合作用的关键因子。寡日照条件下生长的植物Pn低,是由于PAR不足,导致光合作用能量匮乏[4,24]。尤扬等[17]研究表明,在自然环境下生长的黄栌,当PAR达到1 500 μmol/(m2·s)时,其Pn,max为11.1 μmol/(m2·s)。本研究发现,黄栌在生长季遭遇连绵阴雨天气,其叶绿素合成和Pn均受到显著影响,Pn,max仅5.34 μmol/(m2·s),与2018年8月测得同一批黄栌相比[9],2019年8月黄栌Pn,max下降17%,Tr、Rd、Gs下降了48%~66%[9],虽然其光能利用幅度增大了,但对弱光利用能力下降[49-51]。
大量研究表明,植物在逆境条件下,可以通过调节代谢、生长甚至发育来适应环境的变化[18-23,52-54]。李金航等[21]对干旱胁迫下黄栌研究发现,黄栌具有光合生理可塑性,主动降低叶片对光能的吸收,减弱碳同化能力,并根据水资源的变化改变自身对光的获取与利用对策,提高WUE,从而增强其抗旱能力。韩晓等[49]用4种光模型对夏黑葡萄弱光能力分析发现不同组合AQY、LSP、LCP、Rp差异显著,AQY、LSP、LCP、Rp等4个指标很低的组合弱光利用效率低,但耐弱光能力强,呼吸消耗少;AQY、LSP高,LCP、Rp低的组合对光的利用效率高,耐弱光能力强,呼吸消耗少。本研究发现,与大多数阳性植物相比,黄栌具有较宽的光能利用幅度及CO2利用幅度,增强其光合能力及碳同化能力,促进有机物的合成和积累。但在连绵阴雨天气下,光能不足,黄栌叶片AQY仅为2018年正常天气情况下的26%[9],表明其利用弱光能力不强[49-51]。黄栌通过提高耐弱光能力及降低呼吸消耗[50],进而减轻连绵阴雨天气对生长的胁迫作用。
蒸腾作用是植物水分吸收和运输、植物矿物质营养吸收和运输的主要动力,是植物降低和保持体温的重要机制,气孔张开为CO2提供通道,有利于光合作用[14,25,55]。2019年重庆地区连绵阴雨气候下,低光照、高湿度环境不利于气孔开放,空气蒸汽压增大,叶内外蒸汽压差就变小[25],Tr降低的同时,Pn下降、Ci上升,限制光合作用的主要是非气孔因素[50],可能是长时间连绵阴雨天气对黄栌生长发育造成影响,导致光合作用关键酶活性的降低[21,49-51,56-57]。WUE反映植物耗水与干物质生产之间的关系,光照和湿度是其主要影响影子[58],黄栌作为耐干旱树种,通常WUE较高[14,18],与本研究结果一致。
本研究还观察到,遭遇长期阴雨连绵天气的阳性植物黄栌更容易感病,叶片稀疏且容易脱落,姜卫兵等[59]认为这与光照不足、有机物积累减少、生长发育受影响、抗逆能力减弱有关,这些现象有待进一步深入研究。
综合来看,重庆地区连绵的阴雨天气影响黄栌色素合成,黄栌叶片光合效率降低、呈色质量变差。与此同时,黄栌也表现出对不良天气较强的耐受能力,即在低CO2浓度环境具有较宽的生态利用幅度与较宽的光照耐受范围,并通过增加Chlb合成,提高对散射光的利用能力。该研究结果对选育适宜重庆地区寡日照条件下生长的黄栌新品种有一定指导意义。
