斜纹夜蛾取食为害对大豆农艺性状和土壤酶活性的影响
2022-10-13刘建萍李晓红
龙 莹, 刘建萍, 李晓红
(1. 邵阳学院食品与化学工程学院, 邵阳 422000; 2. 邵阳学院城乡建设学院园林系, 邵阳 422000)
大豆Glycinemax,又称黄豆,是一种重要的粮食作物,含有丰富的氨基酸、不饱和脂肪酸、矿物元素和B族维生素等营养成分。在粮食、油料、饲料、工业原料等许多领域中有着广泛的应用。国内外关于大豆品种、种植季节、土壤环境等因素对大豆产量和品质的影响的研究较多,但关于不同密度的昆虫取食为害对大豆产量和营养物质含量的影响的研究相对较少。
植食性昆虫的侵害会严重影响大豆的产量和品质。斜纹夜蛾Spodopteralitura是大豆产区的主要害虫,因其具有暴食性和繁殖快等特点,已成为大豆生长过程中为害最严重的害虫之一[1]。斜纹夜蛾幼虫可取食叶片、花蕾以及果实,严重时可将大豆叶片全部吃光,使其光合作用、物质积累和开花结荚受阻,造成豆粒生长不饱满,豆荚斑痕累累,失去商品价值。近年来我国农作物布局的转变,农业复种指数提高,以及设施农业的大力发展为斜纹夜蛾的连续暴发提供了有利环境。同时由于农药的长期使用,斜纹夜蛾已经产生了抗药性[2]。
植食性昆虫的取食会影响植物的生物量[3]、次生代谢物合成[4]以及根系分泌物的产生等[5]。植物-土壤间存在相互反馈效应。根际吸收土壤中养分提供给植物,植物通过凋零物或根系对土壤进行反馈调控[6]。植物根系分泌物对土壤酸碱度和土壤酶活性存在影响[7]。土壤酸碱度对土壤肥力、微生物活动和作物生长存在影响。土壤脲酶活性可直接反映土壤氮供应的多少,与土壤微生物数量、土壤有机质、全氮和速效氮含量等因素有关。土壤磷酸酶活性影响着土壤养分有效性,能够促进无机磷向有机磷的转化[8]。迄今为止,关于大豆对斜纹夜蛾抗性的研究已有比较深入的研究,但关于斜纹夜蛾取食对大豆土壤酶活性的影响鲜有报道。
植物营养物质对昆虫的生长、发育和繁殖有重要影响,营养物质含量的变化能在一定程度上反映出植物的防御能力[9]。土壤酶活性是反映土壤特征的重要指标,土壤养分可间接影响植物的抗虫能力[10]。本研究随机选用5种常见的大豆品种进行试验,探明不同密度的斜纹夜蛾幼虫取食为害对大豆产量、营养物质含量和土壤酶活性的影响,探究斜纹夜蛾幼虫取食为害对大豆-土壤的相互反馈效应所产生的影响,为筛选抗虫大豆品种及斜纹夜蛾的综合防治提供参考。
1 材料与方法
1.1 材料
供试斜纹夜蛾采自湖南省邵阳市郊区的蔬菜田。采集时为3龄幼虫,随后在人工气候箱内采用人工饲料[11]连续饲养,饲养条件为(26±1)℃、RH(60±10)%和L∥D=14 h∥10 h。
供试大豆品种分别为‘中黄13’‘中黄37’‘中黄39’‘石豆7号’和‘菏豆15’,均来自湖南省种子公司。将供试大豆种子播于营养钵(直径32 cm, 高20 cm)中单株种植,生长期间统一常规管理。
1.2 试验方法
每个品种选取长势基本一致的开花前期大豆进行试验。试验设置4种虫口密度,分别为每株接入0(CK)、3、6、9头斜纹夜蛾2龄幼虫,每处理重复20次,即20株大豆。将斜纹夜蛾2龄幼虫放置到大豆植株中上部的叶片上,用透明薄纱网罩将植株罩住防止幼虫逃逸;每天检查3次,若幼虫死亡或逃逸,及时补充幼虫。幼虫在大豆上取食9 d后,将幼虫和网罩取下。大豆根系外围0~1 cm的土壤视为根际土壤,每株大豆取根际土壤10 g(湿重),将同一处理组的根际土壤混匀,在通风、干燥的环境下自然风干72 h后测定土壤pH(电位法)[12]、脲酶(苯酚钠-次氯酸钠比色法)和磷酸酶(磷酸苯二钠比色法)的活性[13]。同一处理组的土壤pH重复测定10次,每种酶活性重复测定5次。
各处理大豆统一常规管理,观察植株生长发育情况,记录大豆的单株结荚数,大豆成熟后记录单株荚粒数。大豆种子手工脱粒,晒干后测定单株产量。将每处理组收获的大豆种子混匀后,每次随机取50粒粉碎成大豆粉(80目)后测定脂肪含量(GB5009.6-2016索氏抽提法)[14]和蛋白质含量(GB5009.5-2016分光光度法)[15]。每处理重复测定5次。
1.3 数据分析
数据检查方差齐性后,用双因子方差分析法分析不同大豆品种和不同虫口密度斜纹夜蛾取食对大豆农艺性状和土壤酶活性的影响,使用Tukey HSD测验进行多重比较。数据分析使用R软件。
2 结果与分析
2.1 不同虫口密度的斜纹夜蛾取食对大豆单株结荚数的影响
大豆品种、虫口密度以及两者间的互作均对大豆的单株结荚数有显著的影响(大豆品种:F4, 380=156.29,P<0.001; 虫口密度:F3, 380=89.8,P<0.001; 大豆品种×虫口密度:F12, 380=3.45,P<0.001)。在相同虫口密度下,‘中黄39’的单株结荚数显著低于其他4个品种(P<0.05)。不同虫口密度下,各品种大豆的单株结荚数均随虫口密度的增大而降低。‘中黄39’ 的单株结荚数在不同虫口密度处理间差异不显著(P>0.05);‘中黄13’‘中黄37’和‘菏豆15’的单株结荚数在虫口密度≥6头/株时显著低于对照(P<0.05)。在5个品种中,随虫口密度的增加,‘中黄37’的单株结荚数降低最多,当虫口密度为9头/株时,单株结荚数为(28.40±0.61)个(表1),比对照少9.35个(P<0.05)。
2.2 不同虫口密度斜纹夜蛾取食对大豆单株荚粒数的影响
大豆品种、虫口密度以及两者间的互作均对大豆单株荚粒数有显著的影响(大豆品种:F4, 380=133.80,P<0.001; 虫口密度:F3, 380=105.85,P<0.001; 大豆品种×虫口密度:F12, 380=3.25,P<0.001)。在相同虫口密度下,‘中黄13’‘中黄37’和‘菏豆15’的单株荚粒数无显著差异(P>0.05)。不同虫口密度下,各品种大豆的单株荚粒数均随虫口密度的增大而降低。 ‘中黄13’‘中黄37’‘菏豆15’和‘石豆7号’的单株荚粒数在虫口密度≥6头/株时显著低于对照(P<0.05);‘中黄39’的单株荚粒数在虫口密度为9头/株时显著低于对照(P<0.05)。5个品种中,随着虫口密度的增加,‘中黄37’的单株荚粒数降低最多,虫口密度为9头/株时,单株荚粒数为(53.45±1.26)粒(表2),比对照低19.85粒(P<0.05)。
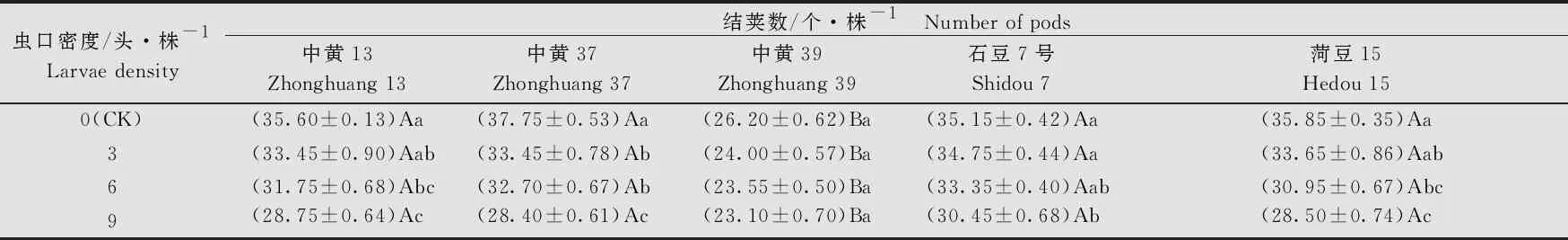
表1 不同虫口密度斜纹夜蛾取食后大豆单株结荚数1)
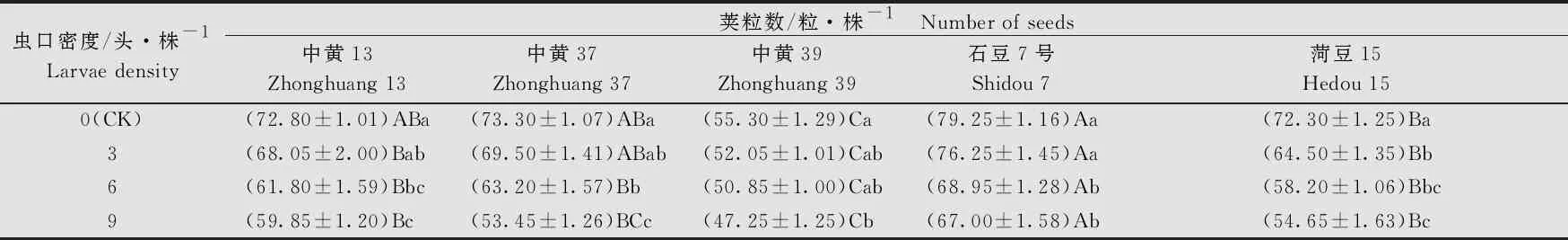
表2 不同虫口密度斜纹夜蛾取食后大豆单株荚粒数
2.3 不同虫口密度斜纹夜蛾取食对大豆单株产量的影响
大豆品种、虫口密度以及两者间的互作均对大豆的单株产量有显著的影响(大豆品种:F4, 380=326.20,P<0.001; 虫口密度:F3, 380=98.89,P<0.001; 大豆品种×虫口密度:F12, 380=1.92,P<0.05)。在相同虫口密度下,‘中黄39’和‘菏豆15’的单株产量均显著低于‘中黄13’‘中黄37’和‘石豆7号’(P<0.05)。不同虫口密度下,各品种大豆的单株产量均随虫口密度的增大而降低,虫口密度为6头/株时,‘中黄13’‘中黄37’‘中黄39’和‘菏豆15’的单株产量与对照相比显著下降(P<0.05);虫口密度为9头/株时,5个大豆品种的单株产量均与对照组差异显著(P<0.05)。在5个品种中,随着虫口密度的增加,‘中黄37’的单株产量减少最多,虫口密度为9头/株时,单株产量为(14.46±0.33)g (表3),比对照低5.18 g(P<0.05)。
2.4 不同虫口密度斜纹夜蛾取食对大豆蛋白质含量的影响
大豆品种、虫口密度以及两者间的互作均对大豆的蛋白质含量有显著的影响(大豆品种:F4, 80=329.53,P<0.001; 虫口密度:F3, 80=75.64,P<0.001; 大豆品种×虫口密度:F12, 80=3.49,P<0.001)。在相同虫口密度下,‘石豆7号’的蛋白质含量均显著低于其他4个品种(P<0.05);在对照组和虫口密度3头/株时,‘中黄13’‘中黄37’和‘菏豆15’间的蛋白质含量无显著差异(P>0.05)。不同虫口密度下,各品种大豆的蛋白质含量均随虫口密度的增大而降低。虫口密度为9头/株时,5个大豆品种的蛋白质含量均显著低于对照(P<0.05)。5个品种中,随着虫口密度的增加,‘中黄13’蛋白质含量减少最多,虫口密度为9头/株时,蛋白质含量为(38.98±0.33)%(表4),比对照低3.36百分点(P<0.05)。
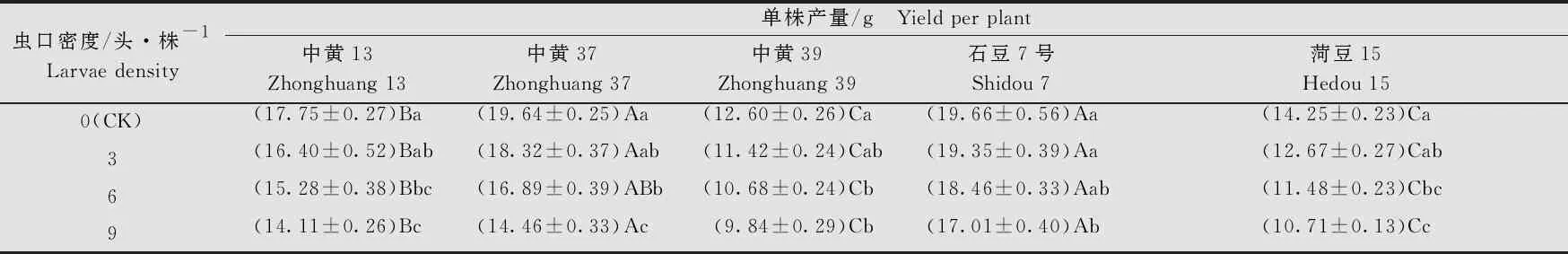
表3 不同虫口密度斜纹夜蛾取食后大豆单株产量
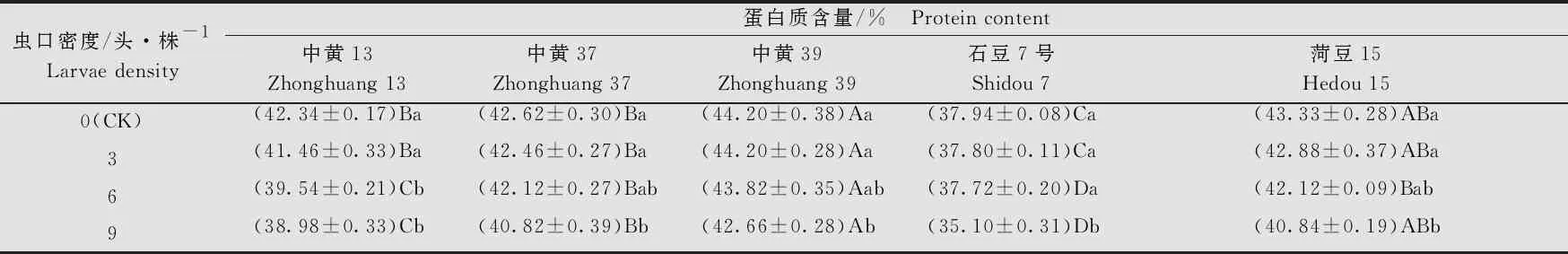
表4 不同虫口密度斜纹夜蛾取食后大豆籽粒中蛋白质含量
2.5 不同虫口密度斜纹夜蛾取食对大豆脂肪含量的影响
大豆品种、虫口密度以及两者间的互作均对大豆的脂肪含量有显著的影响(大豆品种:F4, 80=255.74,P<0.001; 虫口密度:F3, 80=85.40,P<0.001; 大豆品种×虫口密度:F12, 80=5.27,P<0.001)。在相同虫口密度下,‘菏豆15’的脂肪含量均显著低于其他4个品种(P<0.05)。各品种大豆的脂肪含量均随虫口密度的增大而降低,虫口密度为9头/株时,5个大豆品种的脂肪含量均显著低于对照(P<0.05)。其中 ‘中黄37’的脂肪含量在虫口密度为3头/株时与对照相比已表现出显著下降(P<0.05);‘中黄39’和‘菏豆15’ 的脂肪含量在虫口密度为6头、9头/株时显著低于对照(P<0.05);‘中黄13’和‘石豆7号’的脂肪含量在虫口密度为9头/株时显著低于对照(P<0.05)。在5个品种中,随着虫口密度的增加,‘中黄39’的脂肪含量减少最多,虫口密度为9头/株时,脂肪含量为(18.90±0.13)% (表5),比对照低3百分点。
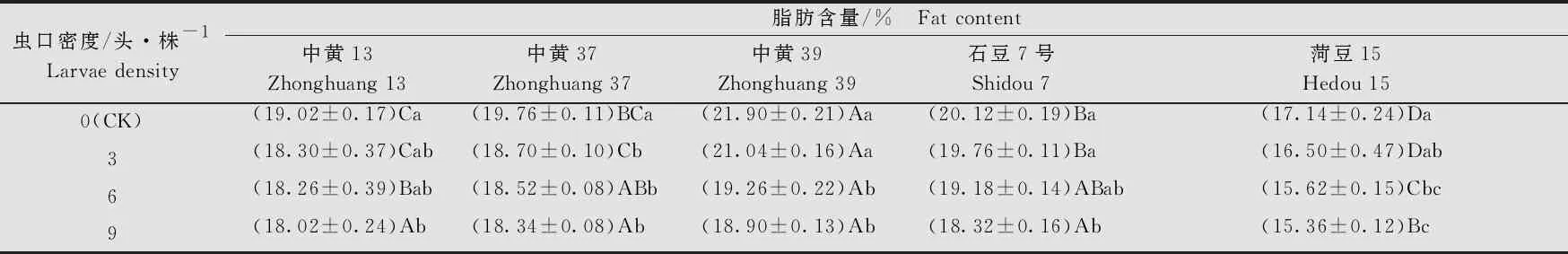
表5 不同虫口密度斜纹夜蛾取食后大豆籽粒中脂肪含量
2.6 不同虫口密度的斜纹夜蛾取食对大豆根际土壤pH的影响
大豆品种、虫口密度以及两者间的互作均对根际土壤pH没有显著的影响(大豆品种:F4, 180=12.57,P>0.05; 虫口密度:F3, 180=6.16,P>0.05; 大豆品种×虫口密度:F12, 180=1.51,P>0.05)。每个品种大豆的根际土壤pH均随虫口密度的增大稍有降低;相同虫口密度下,5个大豆品种间的土壤pH无显著差异(P>0.05)(表6)。

表6 不同虫口密度斜纹夜蛾取食后大豆根际土壤pH
2.7 不同虫口密度的斜纹夜蛾取食对根际土壤脲酶的影响
大豆品种、虫口密度以及两者间的互作均对根际土壤脲酶的活性有显著的影响(大豆品种:F4, 80=10.34,P<0.05; 虫口密度:F3, 80=6.62,P<0.05; 大豆品种×虫口密度:F12, 80=5.89,P<0.05)。在相同虫口密度下,‘石豆7号’分别与‘中黄13’‘中黄37’和‘中黄39’的根际土壤脲酶活性无显著差异(P>0.05)。各品种大豆根际土壤脲酶的活性均随虫口密度的增大而降低。对照组5个大豆品种间的根际土壤脲酶活性均无显著差异(P>0.05);虫口密度为9头/株时,5个大豆品种的根际土壤脲酶活性均显著低于对照(P<0.05)。5个品种中,随虫口密度的增加,‘菏豆15’的根际脲酶活性降低的最多,虫口密度为9头/株时,脲酶活性为(0.095±0.007)mg/(g·d)(表7)。
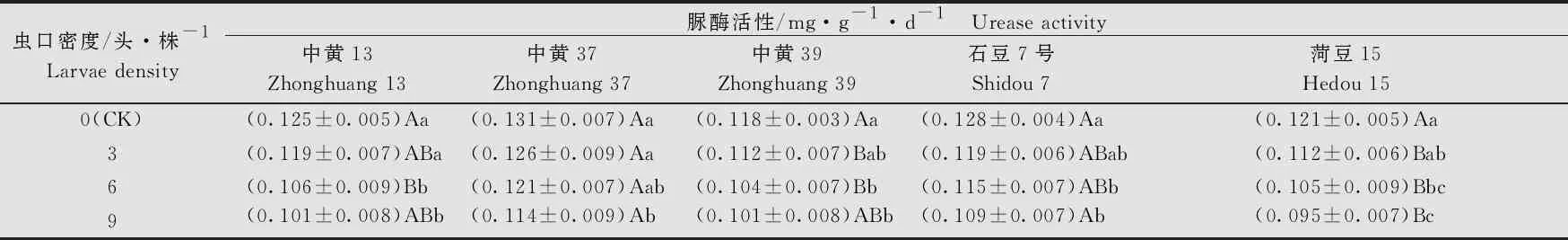
表7 不同虫口密度斜纹夜蛾取食后大豆根际土壤脲酶活性
2.8 不同虫口密度的斜纹夜蛾取食对根际土壤磷酸酶的影响
大豆品种、虫口密度以及两者间的互作均对根际土壤磷酸酶的活性有显著的影响(大豆品种:F4, 80=43.16,P<0.05; 虫口密度:F3, 80=23.87,P<0.05; 大豆品种×虫口密度:F12, 80=14.16,P<0.05)。各品种大豆根际土壤磷酸酶的活性均随虫口密度的增大而降低。在虫口密度≥6头/株时各品种大豆根际土壤磷酸酶活性无显著差异,但均显著低于对照(P<0.05)。在5个品种中,随虫口密度的增加,‘中黄37’的根际磷酸酶活性降低的最多,虫口密度为9头/株时,磷酸酶活性为(2.72±0.14)mg/(g·d)(表8)。
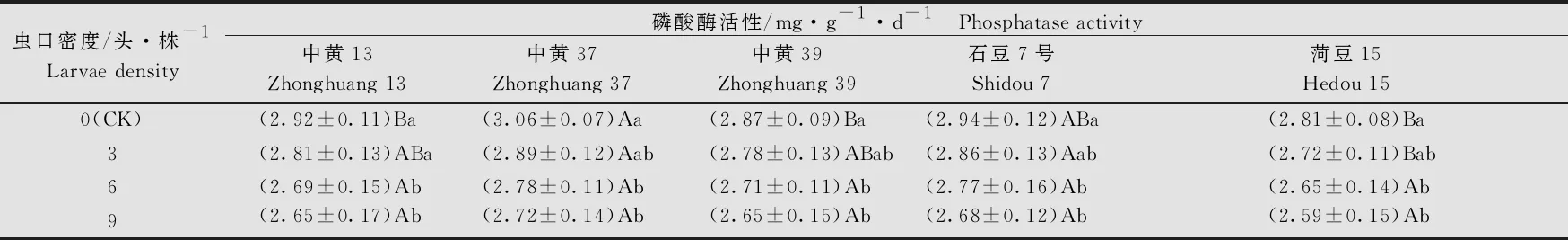
表8 不同虫口密度斜纹夜蛾取食后大豆根际土壤磷酸酶活性
3 结论与讨论
本研究发现斜纹夜蛾取食对不同品种大豆的单株结荚数、单株荚粒数、单株产量、蛋白质含量、脂肪含量以及土壤的脲酶活性、磷酸酶活性均有不利影响,且不利影响均随斜纹夜蛾虫口密度的增加而加重。但虫口密度的变化对土壤酸碱度无显著影响。在5个品种中,虫口密度为9头/株时,‘中黄37’的单株结荚数、单株荚粒数、单株产量和根际磷酸酶活性降低最多,‘中黄13’大豆蛋白质含量降低最多,‘中黄39’大豆脂肪含量降低最多,‘菏豆15’的根际脲酶活性降低最多。
在自然界中,植物受到侵害后,在承受范围内能够自我修复,但超过承受范围时,会导致植物营养物质含量下降或某些次生代谢产物的产生。研究人员发现,斜纹夜蛾的取食损伤了叶片,使叶片的光合能力降低,从而产量降低[16]。豆科植物在营养生长阶段具有较强的补偿性,能较好地弥补昆虫对植物的负面影响而不造成产量损失。然而,当虫害发生在结荚期时,这种补偿是有限的[17]。试验中虫口密度较低时,斜纹夜蛾取食为害后大豆的单株结荚数、单株荚粒数、单株产量、蛋白质含量、脂肪含量与对照无显著差异,原因可能是斜纹夜蛾取食损伤的叶片面积小,对光合作用影响小,植株能够较好地自我修复。但随着虫口密度的升高,叶片残缺面积变大,对光合作用和物质积累的影响超出了植物自身补偿能力范围,因此大豆的结荚数、荚粒数、单株产量、蛋白质含量、脂肪含量显著下降。
在长时间的协同进化历程中,植物形成了抵抗昆虫侵害的防御机制,植物被昆虫为害后,诱导抗性化合物大量形成;而且寄主植物营养物质在质和量上的变化足以影响昆虫的生长发育,因此植物营养物质的改变是植物化学防御的一种对策[9]。本研究中大豆被斜纹夜蛾取食后,蛋白质和脂肪含量均呈下降趋势。易桂林等[18]、尹飞等[19]和蒲晓娟等[20]的研究发现,毛竹被刚竹毒蛾Pantanaphyllostachysae侵害后,蛋白质含量显著减少;芥菜、白菜和芥蓝被小菜蛾Plutellaxylostella为害后,其体内蛋白质含量下降;华山松被华山松大小蠹Dendroctonusarmandi为害后,体内蛋白质、粗脂肪、糖类以及淀粉等营养物质含量均呈下降趋势。这与本研究中大豆蛋白质含量随着虫口密度的增加而下降的研究结果相似,这说明斜纹夜蛾为害阻碍了蛋白质的合成,从而导致蛋白质含量下降。
昆虫取食能够间接改变土壤环境,从而对植物产生影响。土壤酶活性的高低能反映土壤肥力。昆虫取食植物后,植物组织含氮量发生了变化,例如,Sitonaflavescens取食三叶草后,会诱导植物碳氮比显著增加,降低根系含氮量,并改变根系分泌物的组成成分[21]。在松树林中,松白粉蚧Crisicoccuspini和白带梢斑螟Dioryctriaalbovittella的取食作用会显著增加凋落物的氮含量,并且降低其单宁/氮以及碳/氮比例[22]。Classen等发现松白粉蚧对松树的为害使土壤酶活性显著降低[23],这与本研究中土壤酶活性随斜纹夜蛾为害程度加重而逐渐降低的结果相似。
本研究从植物-土壤反馈的生态学角度对斜纹夜蛾-大豆-土壤三者间的相互作用关系进行了探讨,明确了不同密度的斜纹夜蛾幼虫取食会对大豆产量和营养物质含量产生不利影响,对土壤酶活性产生消极作用,并且这一不利影响会随斜纹夜蛾虫口密度的增加而加重。试验结果不仅为了解斜纹夜蛾与大豆之间的关系提供了依据,而且可为斜纹夜蛾在大豆田中的综合防治及大豆抗虫品种的筛选提供参考。
