橡胶树热激蛋白HbHSP90.8-1基因的克隆与功能分析
2022-10-12刘明洋肖化兴王立丰梁晓宇
刘明洋 肖化兴 王立丰 梁晓宇 张 宇 王 萌*
(1. 海南大学植物保护学院,热带农林生物灾害绿色防控教育部重点实验室,海口 570228;2. 农业农村部橡胶树生物学与遗传资源利用重点实验室,省部共建国家重点实验室培育基地—海南省热带作物栽培生理学重点实验室,农业农村部儋州热带作物科学观测实验站,中国热带农业科学院橡胶研究所,海口 571101)
热激蛋白90(heat shock protein 90,HSP90)是一种高度保守和丰富的蛋白,在生物和非生物胁迫下积累,并作为多种蛋白的分子伴侣,其主要参与激酶底物空间结构的形成、底物的激活、初始应激信号传导和维持转录因子空间结构等。例如,当HSP90 伴侣活性较高时,生长素受体蛋白TIR1 正确折叠形成功能性SCF复合体,对促使Aux/IAA 转录因子降解起关键作用。HSP90 被蛋白激酶HopBF1 磷酸化后会使ATP 酶活性和伴侣功能失活,导致触发植物过敏反应免疫受体的激活受阻止。在植物中,对病原菌最有效的特异性抗性来自抗病R 基因,R 基因大多数编码NLR蛋白,HSP90 通过调节植物NLR 蛋白的活性来应对病原菌的侵染。植物中如果没有HSP90 蛋白会降低拟南芥()RAR1 对R 基因介导的免疫功能,同时也会降低免疫受体MLA1和MLA6 的积累,表明HSP90 对NLR 的稳定性和免疫受体复合体的形成至关重要。HSP90的分子伴侣SGT1 是维持Rx 和N 在内的NLR 的稳态水平所必需。SGT1 不仅在调节植物NLR 活性方面发挥积极作用,而且在控制NLR 蛋白如RPM1、RPS5 和SNC1 的循环方面也发挥消极作用。研究证明HSP90 与分子伴侣等底物多肽相结合来调控植物抗病性。在大麦()中HSP90-RAR1-SGT1 相互作用提高对大麦条锈病的抗病性。在番茄()中HSP90与SGT1相互作用提高番茄抗性基因Tm-22对TMV 的抗病功能。并且已有研究表明基因沉默还会引起泛素化蛋白积累和细胞死亡,对番茄黄化曲叶病毒有缓解作用。HSP90 除了在植物免疫应答中发挥积极作用外,HSP90 还参与了NLR 的周转控制,这种调控对于维持适当水平的NLR 蛋白至关重要,以避免可能导致自身免疫缺陷的免疫受体的过度积累。
橡胶树()是热带地区典型的经济树种,所产的天然橡胶在军工、航天及医疗等具有特殊和重要的地位。橡胶树在生长发育过程中面临许多病害侵染,造成橡胶产量损失,橡胶树白粉病()是橡胶树主要叶部病害之一。白粉病主要危害橡胶树的幼嫩组织,发病严重时,病叶布满白粉,皱缩畸形,最后脱落,从而造成胶乳产量降低。近年来,随着基因在植物中的作用机制深入的研究、分子生物学等新技术的高速发展以及橡胶树遗传转化技术体系的建立,培育橡胶树白粉病抗病品种是有效且经济的手段。目前,关于基因在橡胶树中的功能还鲜有报道,揭示其在橡胶树中的特异性功能有助于拓宽对的认知。为解析在橡胶树中的抗白粉病功能,本研究利用前期橡胶树基因组数据,通过PCR 技术克隆得到-基因,利用生物信息学对其结构和功能进行分析;同时通过荧光定量对其在白粉菌侵染、植物抗病相关激素及HO处理中表达量分析,为深入研究-基因功能及调控橡胶树抗白粉病分子机制奠定基础。
1 材料与方法
1.1 试验材料
于中国热带农业科学院橡胶研究所国家橡胶树种质资源圃选取健康10年树龄正常割胶的橡胶树品种热研73397 的根、花、枝、茎、叶和胶乳为材料,用于-基因的克隆和在不同组织中的表达分析。选取株高70~80 cm橡胶树热研73397组培苗用于白粉菌侵染、HO和不同激素等处理。
植物RNA 提取试剂盒(货号:DP441)购自天根生化科技有限公司,反转录试剂盒、大肠杆菌()DH5-感受态、农杆菌GV3101菌株购自TaKaRa 公司。PCR 引物由上海铂尚生物技术有限公司合成。Phanta 高保真酶、ClonEXpress Ⅱ试剂盒、第二代TOPO 克隆试剂盒均购自南京诺唯赞公司。
1.2 试验方法
橡胶树品种热研73397 的根、茎、叶、花、枝和胶乳作为-基因克隆和不同组织的表达量分析。在橡胶树品种热研73397 组培苗上分别喷施200 μmol·L水杨酸(Salicylic Acid,SA)、200 μmol·L茉莉酸甲酯(Methyl Jasmonate,Me-JA)、200 μmol·L脱落酸(Abscisic Acid,ABA)、1%(V/V)乙烯(Ethylene,ETH)、2%(V/V)过氧化氢(Hydrogen Peroxide,HO),对照组植株喷施0.05%(V/V)的乙醇水溶液。在喷施激素后的0、0.5、2、6、10、24 h 采集试验组材料,对照组同时采样。白粉菌侵染试验前,对橡胶树品种热研73397隔离一周后再进行白粉菌侵染试验,采集侵染后0、3、6、12、24、48、72、96 h 的叶片,对照组同步采样。样品均用液氮速冻后放入冰箱中-80 ℃保存,每样品3次生物学重复。
试验前使用75%乙醇清洁实验区和灼烧研钵、勺子等器具,并将液氮分装于保温壶中备用。采集的所有样品参考天根生化科技有限公司的RNAprep Pure植物总RNA提取试剂盒说明书步骤进行提取,采用Thermo Fisher NanoDrop 2000 超微量核酸蛋白分析仪检测总RNA 浓度和纯度,提取后的RNA 于冰箱中-80 ℃保存备用。RNA 反转录方法按照TaKaRa 反转录试剂盒实验步骤,以橡胶树内参基因HbActin-F/R 为引物检测cDNA 质量,并以1.0%琼脂糖凝胶电泳检测RNA 完整性及cDNA的浓度和纯度。
根据橡胶树基因组获得-基因序列,利用Primer premier 6.0软件设计引物--F,5′-AGAAGCTGCTAGAAACCG-3′,--R,5′-TTCCTGTCATACGCTGAAC-3′,引物序列由上海铂尚生物技术有限公司合成。以反转录得到的橡胶树热研73397 叶片的cDNA 为模板,按照高保真酶试剂盒说明书,利用PCR 扩增橡胶树-基因的全长序列,将目的条带按胶回收试剂盒进行回收纯化,目的片段连接PMD-18T 载体,水浴16 ℃连接4 h,转化大肠杆菌DH5α 感受态细胞中,经PCR检测后挑取阳性克隆测序。
利用在线分析软件对-进行生物信息学预测分析(见表1)。利用DNAMAN 工具软件进行多序列比对。MEGAX 构建系统进化树(neighbor-joining,bootstrap 值设为1 000)。用于系统进化分析的蛋白序列从PlantTFDB、NCBI下载。

表1 生物信息学在线分析工具Table 1 Bioinformatics online analysis tool
根据-基因序列ORF 区设计实时荧 光 定 量 引 物--QF,5′-GGGCCTAAAACTTGGGAAAG-3′;--QR,5′-GACGTCACAACCACACAAGG-3′以反转录的cDNA为模板,橡胶树(GenBank:HQ260674.1)基因为内参,荧光定量引物-F,5′-GATGTGGATATCAGGAAGGA-3′;-R,5′-CATACTGCTTGGAGCAAGA-3′;荧光定量采用SYBR Green 法进行qRT-PCR 扩增。通过qRT-PCR 技术分析-基因在白粉菌侵染、植物抗病相关激素、HO及不同组织处理的表达模式。
基因表达量为3 次生物学重复和3 次技术重复的平均值±标准误。基因相对表达结果在Excel 2016 软 件 中 用2法进 行 分 析。使 用IBM SPSS Statistics 25 软件进行统计分析,采用单因素ANOVA 检验分析差异显著性,使用Origin 2018作图。
2 结果与分析
2.1 HbHSP90.8-1基因的克隆
以橡胶树热研73397 健康树叶片的cDNA 为模板,采用PCR 扩增-,结果如图1 所示,扩增出了与预期大小相符的特异性条带。测序验证后,将cDNA序列命名为-(Gen-Bank:MW413357)。-基因的cDNA 全长序列2 844 bp,包含2 454 bp 的开放阅读框(ORF),编码817个氨基酸。

图1 HbHSP90.8-1基因扩增结果M.DL5000 DNA Marker;1.目的基因扩增产物Fig.1 The PCR amplification products of HbHSP90.8-1M.DL5000 DNA Marker;1.The objective gene amplification products
2.2 HbHSP90.8-1基因生物信息学分析
利用ProtParam 工具在线分析HbHSP90.8-1氨基酸编码蛋白质的理化性质:分子式CHNOS,总原子数13 118,分子质量为93 526.92 Da,等电点4.88,总平均亲水性为-0.757,带正电残基总数(Arg+Lys)为121,带负电荷残基总数(Asp+Glu)为163,不稳定系数为34.33(该蛋白属于稳定蛋白),脂肪族氨基酸指数是76.38,推测HbHSP90.8-1 蛋白是一个稳定的亲水蛋白。采用TMHMM Server v.2.0 在线工具分析预测表明HbHSP90.8-1蛋白无跨膜结构。NCBI保守结构域工具分析预测表明HbHSP90.8-1 蛋白存在HATPase-Hsp90-like 和HSP90 superfamily 结构域,其位置分别位于87~277 和259~780 aa 处(见图2A)。利用SOPMA 分析HbHSP90.8-1 蛋白的二级结构,结果显示螺旋占50.67%,延长链占12.73%,β 折叠占4.65%,无规卷曲占31.95%(见图2B)。利用SWISS-MODEL 分析HbHSP90.8-1 的三级结构如图2C 所示。利用DNAMAN 软件对来源于不同植物的HSP90 相关氨基酸序列进行同源比对(见图3A),结果发现橡胶树HbHSP90.8-1 与木薯(,XP_021601014.1)、麻风树(,KDP36908.1)、蓖麻(,XP_002510550.1)等不同物种的HSP90 蛋白均具有较高的同源性。多序列比对分析并根据图2A 分析的结果标注HbHSP90.8-1 保守结构域(见图3A)。采用MEME 工具分析橡胶树HbHSP90.8-1与其在NCBI数据库中比对亲缘性较高的HSP90 蛋白序列,预测了排名前三的motif(见图3B)。为了分析橡胶树HbHSP90.8-1 与其他植物HSP90 蛋白的系统进化关系,利用MEGA-X 软件构建系统进化树,结果表明橡胶树HbHSP90.8-1与木薯MeHSP90 的亲缘关系最近,在分类上与麻风树JcHSP90 聚为一类,这预示着他们的功能可能具有相似性(见图4A)。SignalP-5.0 Server 分析预测HbHSP90.8-1 存在信号肽的概率为98.5%。DeepLoc1.0 预测分析HbHSP90.8-1蛋白定位于内质网上(见图4B)。综上,HbHSP90.8-1可编码一个分泌型且无跨膜结构的蛋白。
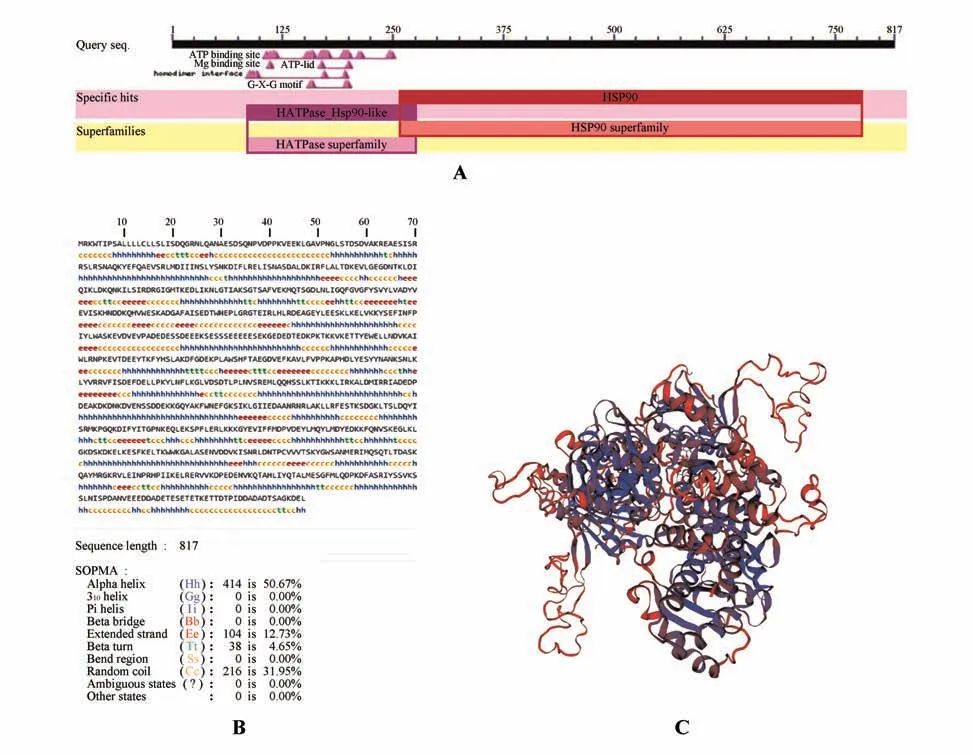
图2 HbHSP90.8-1结构分析A.保守结构域预测;B.二级结构预测;C.三级结构预测Fig.2 Structural analysis of HbHSP90.8-1A.Prediction of the conserved domains;B.Prediction for the secondary structure;C.Prediction for the tertiary structure

图3 HbHSP90.8-1与其他植物HSP90蛋白序列A.HbHSP90.8-1与其他植物HSP90蛋白序列多重对比;B.HbHSP90.8-1与其他植物HSP90蛋白序列MEME聚类Fig.3 Protein sequence analysis of HbHSP90.8-1 with other plant HSP90 proteinsA.Multiple protein sequence alignment of HbHSP90.8-1 with other plant HSP90 proteins;B.MEME cluster analysis of HbHSP90.8-1 with other plant HSP90 proteins.
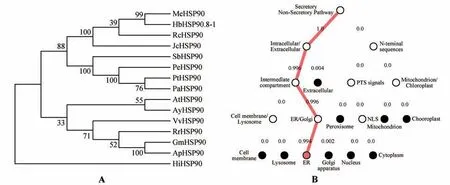
图4 HbHSP90.8-1的聚类分析和亚细胞定位分析A.橡胶树HbHSP90.8-1 蛋白与其他物种HSP90 蛋白序列的系统进化树分析(大豆GmHSP90(Glycine max,XP_003545030.1);相思豆ApHSP90(Abrus precatorius,XP_027368246.1);银白杨PaHSP90(Populus alba,XP_034931576.1);红脉麦果RrHSP90(Rhamnella rubrinervis,KAF3446793.1);葡萄VvHSP90(Vitis vinifera,XP_002273785.1);紫花风铃木HiHSP90(Handroanthus impetiginosus,PIN15193.1);拟南芥AtHSP90(Arabidopsis thaliana,NP_194150.1);漾 濞 槭AyHSP90(Acer yangbiense,TXG52042.1);小 垫 柳SbHSP90(Salix brachista,KAB5557693.1);胡杨PeHSP90(Populus euphratica,XP_011045067.1);毛果杨PtHSP90(Populus trichocarpa,XP_002307732.1);蓖麻RcHSP90(Ricinus communis,XP_002510550.1);木 薯MeHSP90(Manihot esculenta,XP_021601014.1);麻 风 树JcHSP90(Jatropha curcas,KDP36908.1),橡胶树HbHSP90.8-1);B.HbHSP90.8-1亚细胞定位预测Fig.4 Phylogenetic tree analysis and subcellular localization prediction of HbHSP90.8-1A.Phylogenetic tree analysis of HbHSP90.8-1 protein with other species HSP90 proteins;GmHSP90(Glycine max,XP_003545030.1);ApHSP90(Abrus precatorius,XP_027368246.1);PaHSP90(Populus alba,XP_034931576.1);RrHSP90(Rhamnella rubrinervis,KAF3446793.1);VvHSP90(Vitis vinifera,XP_002273785.1);HiHSP90(Handroanthus impetiginosus,PIN15193.1);AtHSP90(Arabidopsis thaliana,NP_194150.1);AyHSP90(Acer yangbiense,TXG52042.1);SbHSP90(Salix brachista,KAB5557693.1);PeHSP90(Populus euphratica,XP_011045067.1);PtHSP90(Populus trichocarpa,XP_002307732.1);RcHSP90(Ricinus communis,XP_002510550.1);MeHSP90(Manihot esculenta,XP_021601014.1);JcHSP90(Jatropha curcas,KDP36908.1);HbHSP90.8-1B.Prediction of HbHSP90.8-1 subcellular localization
2.3 HbHSP90.8-1表达模式分析
通过实时荧光定量PCR 数据发现,-在橡胶树不同组织中的表达量差异显著(见图5A)。-在橡胶树的根、花、枝、茎、叶胶、乳中均有表达,其中在胶乳中的表达量较高,枝次之,在根、茎、叶的表达量较低,胶乳的表达量是根、花、叶的28倍左右,表明-基因主要作用在胶乳中。白粉菌侵染橡胶树叶片后,-基因的表达量呈现上调趋势,在6 h表达量达到最高,是处理前的4倍,表明白粉菌侵染会导致-基因表达量显著性上升(见图5B)。在HO处理下,橡胶树-基因的表达量均显著上调,在处理后的10 h 表达量达到处理前的4.5 倍,表明-基因受HO的诱导上调表达(见图5C)。在ABA和ETH激素处理下,-基因表达量呈现先上升的表达趋势,分别在2、10 h表达量达到最高,分别是处理前的26、21倍,表明-基因受ABA 和ETH激素诱导上调表达(见图6:A、B)。在MeJA和SA激素喷施处理后,-基因在橡胶树中的表达量如图所示(见图6:C、D)。-基因的表达量呈现下调的趋势,-基因在MeJA处理和SA处理的2 h表达量达到最低,分别是处理前的8.5、14.0倍,表明JA和SA激素的处理会导致-基因表达量的显著下降。

图5 HbHSP90.8-1 基因在组织、白粉菌侵染和H2O2处理叶片表达模式A.组织;B.白粉菌侵染处理;C.H2O2处理Fig.5 Expression pattern of HbHSP90.8-1 gene in tissue,drought,powdery mildew infection,H2O2,mechanical wounding treatmentA.Tissue;B.Oidium heveae infection;C.H2O2 treatment
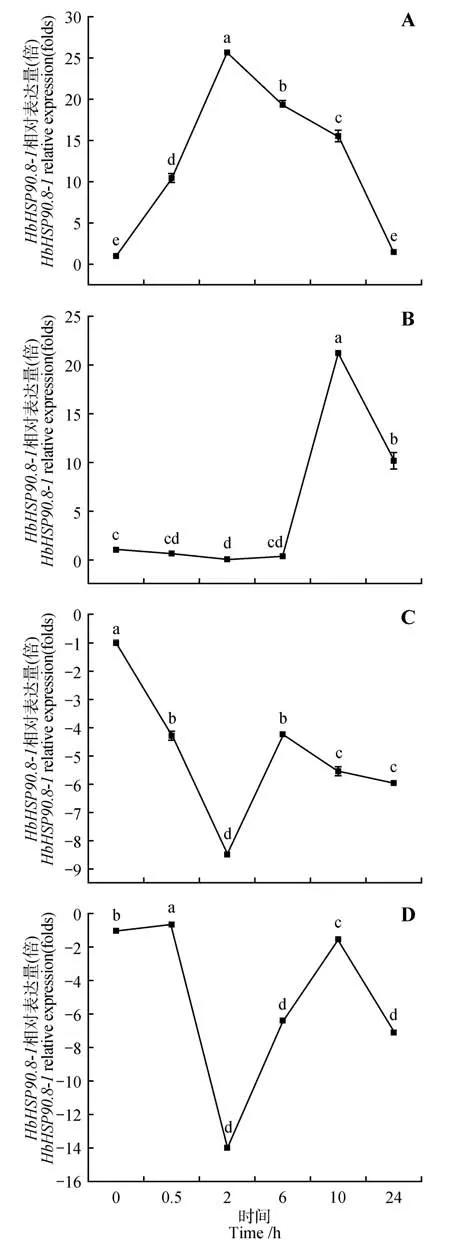
图6 HbHSP90.8-1基因在不同激素处理的表达模式A.ABA激素处理;B.ETH激素处理;C.MeJA激素处理;D.SA激素处理Fig.6 Expression pattern of HbHSP90.8-1 gene in different hormone treatmentsA.ABAtreatment;B.ETHtreatment;C.MeJAtreatment;D.SAtreatment
3 讨论
植物在生长过程中会受到各种逆境胁迫的影响,包括高温、高盐和病原菌侵染等胁迫。热激蛋白HSP90 在植物抗病过程及稳定和激活各种关键信号蛋白方面发挥重要作用。为了解-基因在橡胶树中的抗白粉病功能,本研究从橡胶树品种热研73397 的叶片克隆了-基因。生物信息学分析发现,HbHSP90.8-1存在HATPase-Hsp90-like和HSP90 superfamily结构域。ATP结构域是ATP/ADP 结合位点,具有内源ATP 活性。ATP 存在情况下会参与细胞间信号转导和多肽折叠等。且有研究表明HSP90的ATPase 结构域与RAR1 的CORDI 域结合,同时与SGT1 蛋白的CS 域特异结合,对于拟南芥RPS2介导的抗病性起核心作用。推测-基因参与橡胶树抗病响应过程。HbHSP90.8-1 定位预测在内质网上,是一个有信号肽、无跨膜结构的稳定亲水蛋白。在杨树基因家族成员通过亚细胞定位结果表明-和-分别定位在叶绿体和线粒体上,-定位在内质网中,在应激情况下,HSP90s 会进入细胞核和核仁发挥作用。推测-在参与橡胶树抗白粉病时可能会迁移到细胞核中发挥作用。
本研究通过qRT-PCR 分析-表达模式,结果表明-在橡胶树根、花、枝、茎、叶、胶乳中均有表达,但主要在橡胶树的胶乳中表达。番茄和辣椒中的在绝大多数组织中也都有表达。表明-与大部分基因的表达特性一致。R 基因编码的NBLRR 蛋白参与感知病原体产物和调节细胞信号和死亡。MLA 属于NB-LRR 蛋白之一,参与植物的抗病过程。大麦中MLA基因对白粉病的抗性是通过识别SGT1-Rar1 复合体在通路的下游对白粉病起抗病作用。研究表明基因通过VIGS方法沉默后发现介导的大麦白粉菌抗性减弱,证明了基因通过调控基因参与对白粉病的抗性过程。HSP90的分子伴侣SGT1也参与橡胶树白粉病的抗病性。在橡胶树叶片上接种白粉菌后,通过观察菌丝生长状态,发现突变体表现出稀疏的菌丝网络,并且突变体上活性氧的产生也降低了,表明参与橡胶树白粉病的抗病反应。推测HSP90 与分子伴侣SGT1 相互作用,从而启动一个特异性信号级来参与抗病反应过程。在本研究中接种白粉菌后发现,-基因的表达量呈现上调趋势,在6 h表达量达到最高,是处理前的4倍,表明白粉菌侵染会导致-基因表达量显著性上升。与的 研 究 结 果 一 致。推 测-基因可能参与橡胶树白粉菌抗性过程。SA、JA、ETH、ABA 等多种植物激素调控的信号网络相互作用对植物抗病能力有重要影响。已有研究发现经SA、MeJA 和ETH 处理显著上调了南瓜抗白粉病自交系中的表达,表明响应植物激素信号途径。在南瓜中采用qRTPCR 分析显示基因对SA 处理具有明显应答反应,说明基因也响应SA 激素处理。ETH 与ABA 在EIN3-AB14-VTC2 级联蛋白在抗坏血酸AsA(ascorbic acid)生物光合过程调控活性氧ROS(reactive oxygen species)积累以应对病原菌对植物的侵染。ABA 和ETH 处理后发现,-基因表达量显著上调表达,分别是处理前的26、21 倍,在MeJA 和SA 处理后发现,-基因的表达量呈现显著下调的趋势,在2 h 表达量达到最低,分别是处理前的8.5、14 倍,表明-响应ABA、ETH、JA 和SA 信号途径,推测ABA、ETH、JA 和SA 作为信号分子参与-抗病防御反应过程中。植物在受到病原菌感染后,会产生过敏性反应,感染部位会有大量活性氧ROS产生和积累。白粉菌侵染会造成叶片组织结构损伤,使植物产生过敏反应来应对白粉病原菌。在HO处理下,发现橡胶树-基因上调表达,推测-基因参与植物体内HO诱导抗病传导过程。综上所述,初步认为-基因不仅响应白粉菌侵染胁迫的应答,还受植物抗病相关激素和HO诱导,推测-在橡胶树白粉病抗病传导过程及激活植物免疫过程中有重要作用。以上研究结果为进一步了解橡胶树-基因的功能以及橡胶树抗白粉病分子作用机制奠定基础。
