黎母山引进外来树种对森林群落物种多样性的影响
2022-10-04方发之黎肇家桂慧颖
方发之,黎肇家,桂慧颖
(海南省林业科学研究院/海南省红树林研究院,海南 海口 571100)
外来树种是一定区域内历史上没有自然分布,通过人为活动直接或间接引入的树种[1]。多年以来,为促进农林业发展,我国大量引入外来树种丰富特定区域生物多样性和提高农林业经济效益[2-3]。尽管外来树种发展为有害植物的可能性比较大,但在此之前并不一定对当地生态系统造成威胁[1]。外来树种在新的环境内往往具有较高生长率和较强环境适应能力[4],生长速度快,在山体损坏区域种植,能快速形成林分减少水土流失,对山体生态修复具有积极作用。对破损山体生态修复优先发展乡土树种是有利于形成较为稳定的生态系统,但乡土树种生长速度慢,在生态脆弱地区修复不适合早期种植,应由速生树种生长为优势树种之后再引入乡土树种,既能在短时间内保护生态,又能让乡土树种回归大自然,促进生态安全[2,5]。
加勒比松(Pinuscaribaea)分布于加勒比海地区,1964年我国引种,1974年海南引种,为速生用材树种[6-7],在海南多地有种植,属于海南热带雨林国家公园外来树种。马占相思(Acaciamangium)为豆科金合欢属(Acacia)乔木树种,原产于澳大利亚,1979 年引入我国,同年引入海南,具有速生丰产[8]、固氮改良土壤作用[9],能提供高效的经济和生态效益,属于海南热带雨林国家公园外来树种之一。橡胶树(Heveabrasiliensis)原产于亚马逊流域,对水分和热量要求比较高[10],1904年我国引种,在海南多地有种植,是海南重要化工原料植物[11],属于海南热带雨林国家公园外来树种。桉树(Eucalyptusspp.)是桃金娘科桉属树种总称[12],海南1917年前后引种,20世纪80年代,桉树得到大量发展机会,是重要经济用材树种[13],属于海南热带雨林国家公园外来树种之一。
本研究以海南热带雨林国家公园黎母山片区内4个外来树种和1个次生林森林群落植被为对象,通过调查4个外来树种森林群落植被物种多样性,探究外来树种森林群落植被物种多样性趋势,为外来树种人工林群落修复技术提供数据参考。
1 材料与方法
1.1 研究区概况
研究区为海南热带雨林国家公园,地处海南省,总面积约4 403 km2,地理坐标108°44′32″-110°04′43″E,18°33′16″-19°14′16″N。海南热带雨林国家公园属于典型热带季风气候,年平均气温22.5 ℃,年均降水量2 343.1 mm,全年高温多雨[14]。由黎母山、尖峰岭、霸王岭、吊罗山等片区组成,其中黎母山片区位于海南岛中部,琼中、白沙两县境内(109°39′05″-109°48′31″E,19°07′22″-19°14′03″N),总面积约554.87 km2,占海南热带雨林国家公园总面积的12.61%。黎母山片区人工林面积约186.67 km2,占黎母山片区总面积的33.64%,其中桉树人工林面积约50.01 km2,占人工林面积的26.79%;加勒比松人工林面积约52.52 km2,占人工林面积的28.13%;相思类人工林面积约27.14 km2,占人工林面积的14.54%;橡胶人工林面积约37.77 km2,占人工林面积的20.23%。黎母山主峰海拔1 412 m,林区大部分土壤为花岗岩和砂岩风化而成的砂壤土和少部分轻壤土。土壤类型以黄壤、赤红壤和砖红壤为主,为偏酸性土壤[14-15]。海南热带雨林国家公园有丰富的天然热带雨林和大面积的人工纯林资源,林区内生长着多种国家重点保护植物,植被资源丰富,阳光充足。
1.2 研究方法
以具有代表性的4个外来树种不同树龄森林群落和1个次生林森林群落为调查区域。外来树种森林群落建群树种为人工营造外来种,本研究对象为人工引种后由林地内生长起来的其他乔灌草植被群落(不含建群树种,所描述乔木或乔木层均不含建群树种);次生林森林群落优势树种有贡甲(Maclurodendronoligophlebium)、青梅(Vaticamangachapoi)等,本研究对象为人工采伐后自然更新形成的乔灌草植被群落(含优势树种)。调查森林群落分别为1993年种植加勒比松人工林群落(J-28)、2006种植年加勒比松人工林群落(J-15)、2001年种植马占相思人工林群落(M-20)、1995年种植马占相思人工林群落(M-26)、2005年种植橡胶人工林群落(X-16)、2000年种植橡胶人工林群落(X-21)和次生林森林群落(CK),其中次生林森林群落为对照(CK)。调查于2021年3月进行,样地信息见表1。
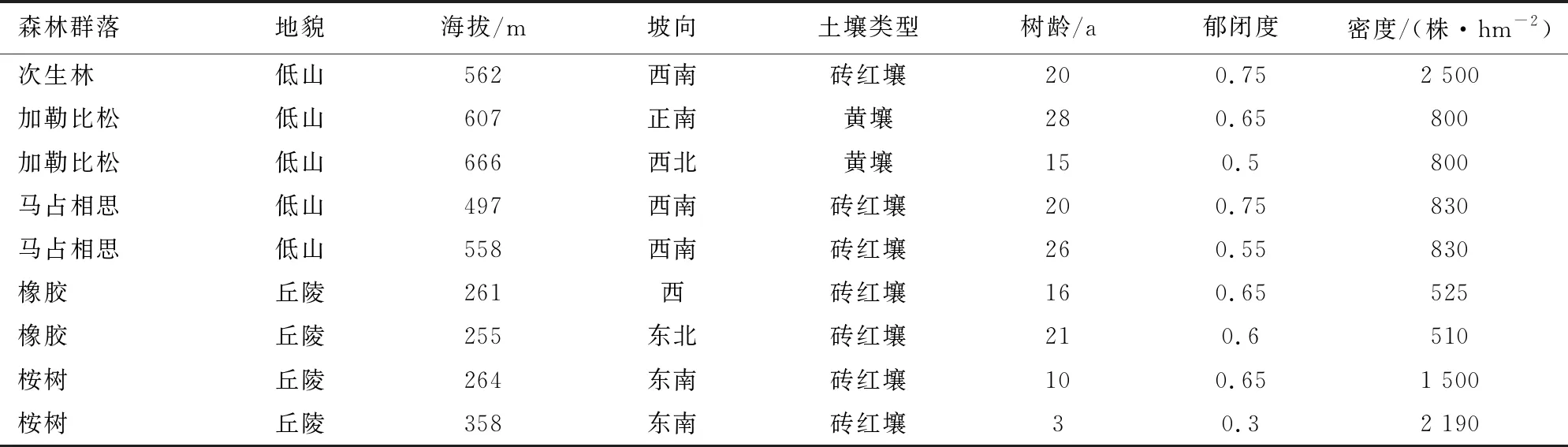
表1 样地基本情况
采用样方法调查,在林分内布设3个20 m×20 m调查样方,对样方内乔木层所有树种进行每木检尺,记录种名、胸径、树高、冠幅等,胸径小于2 cm的幼树记录树种名称;每个乔木层样方4个角布设2 m×2 m调查样方为灌木层调查区域,草本层每个样方布设4个1 m×1 m调查样方,对灌木层和草本层植物进行调查,记录种名、高度、盖度、株数或丛数。
1.3 树种重要值及物种多样性计算
根据外业调查数据结果,内业计算相关植物种类重要值(important value,IV)。乔木层重要值=(相对密度+相对高度+相对显著度)/3;灌木层和草本层重要值=(相对密度+相对高度+相对盖度)/3。并在此基础上计算各个群落物种多样性指数,采用Shannon-Wiener指数H′,Simpson指数H、Pielou均匀度指数Jsw综合评价各个调查群落物种多样性。
相对显著度C:
(1)
Simpson优势度指数H:
(2)
Shannon-Wiener多样性指数H′:
(3)
Pielou均匀度指数Jsw:
(4)
式中:d为某个树种胸径断面积和,D为所有树种胸径断面积和,Pi为第i种的个体数ni占所有种个体总数n的比例,ni为第i种的个体数,n为所有种个体总数,即Pi=ni/n;i=1,2,3,…S,S为物种数。
2 结果与分析
2.1 群落各层次物种数量
由表2可知,CK乔木层、灌木层和草本层的物种数分别为40、3和0种。J-28和J-15乔木层、灌木层和草本层的物种数分别为21、4、2种和12、3、2种,J-15乔木层物种数比J-28少42.86%。M-20和M-26乔木层、灌木层、草本层的物种数分别为29、5、2种和9、1、3种。X-16和X-21乔木层、灌木层、草本层的物种数分别为8、4、3种和0、0、2种。A-10和A-3乔木层、灌木层和草本层的物种数分别为8、4、2种和0、2、7种,10 a树龄桉树已经有利于乔木树种生长。
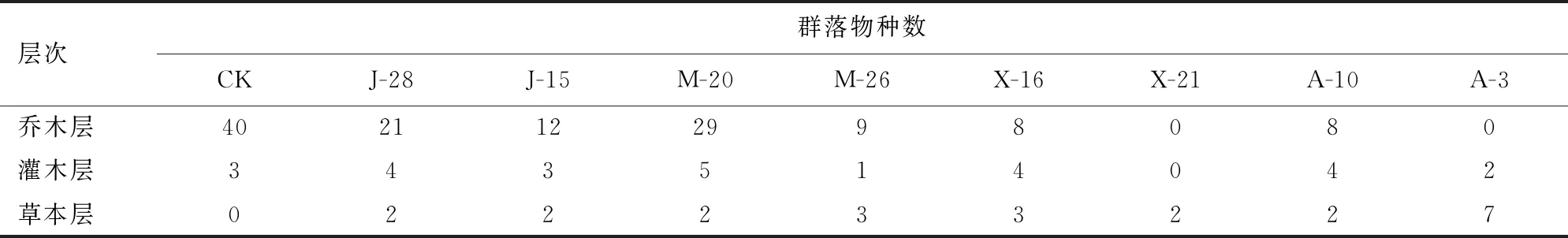
表2 调查样地森林群落各层次物种数
2.2 重要值分析
2.2.1 森林群落乔木层树种重要值 由表3可知,CK乔木层主要优势树种有贡甲和青梅,其中贡甲重要值17.87%为该群落乔木层树种重要值最大,其次为青梅14.76%,密花树(Rapaneaneriifolia)重要值最低为0.49%。J-28乔木层主要优势树种为鸭脚木,重要值为38.94%,其次为水锦树(Wendlandiauvariifolia)10.22%,最小为禾串树(Brideliabalansae)0.94%,优势树种十分明显。J-15乔木层主要优势树种有中平树(Macarangadenticulata)和水同木(Ficusfistulosa),重要值分别为37.99%和24.25%,其余树种重要值分布比较均匀。M-20乔木层树种重要值最大为细齿叶柃(Euryanitida)18.80%,其次为黄牛木(Cratoxylumcochinchinense)重要值18.37%,其余树种重要值相对偏低。M-26乔木层以八角枫(Alangiumchinense)重要值最大,为30.68%,鸭脚木重要值18.78%,青藤公(Ficuslangkokensis)重要值14.94%,其余树种重要值均小于10%,与M-20相比较,该群落乔木层树种比较少。X-16乔木层白楸(Mallotuspaniculatus)重要值最大,芳槁润楠(Machilusgamblei)最小,分别为31.64%和1.61%,其余树种重要值相差明显,优势树种白楸相对突出,原因是前10 a该群落人为管护程度相对高,到后6 a人为管护程度小,导致白楸等速生杂生树种快速繁殖, X-21乔木层树种生长受人为干扰程度高,乔木难以获得正常生长机会。A-10乔木层细基丸(Polyalthiacerasoides)重要值最大,为45.74%,倒吊笔(Wrightiapubescens)重要值最小,仅2.57%,同时A-3没有乔木。

表3 调查样地群落乔木层树种重要值

续表3
加勒比松、马占相思和橡胶树乔木相同树种比较多,桉树群落乔木树种与另3个外来树种群落乔木相同树种比较少,可能是由于桉树地理偏离,同时也更有利于桉树林群落灌木和草本植物的生长。
2.2.2 群落灌木层植物重要值 由表4可知,香楠为CK灌木层优势灌木树种,重要值50.60%。毛菍为J-28灌木层优势树种,重要值为46.72%。山柑算盘子为J-15灌木层优势树种,重要值为36.67%,毛菍重要值为32.70%,豆腐柴重要值为30.63%。M-20和M-26灌木树种相差较大,M-26只有荚蒾(Viburnumdilatatum)一种灌木,更新能力比较差;M-20优势树为黑面神,其重要值为23.25%。X-21灌木树种数量为0,受人为管护程度比较大,灌木树种被清除,同时X-16受管护程度变化使灌木层部分树种得到生长机会,其中山椒子和多花野牡丹的重要值分别为37.26%和31.37%,2个树种具有显著优势。A-10和A-3灌木层优势树种分别为大青,重要值48.84%,海南狗牙花重要值59.83%。
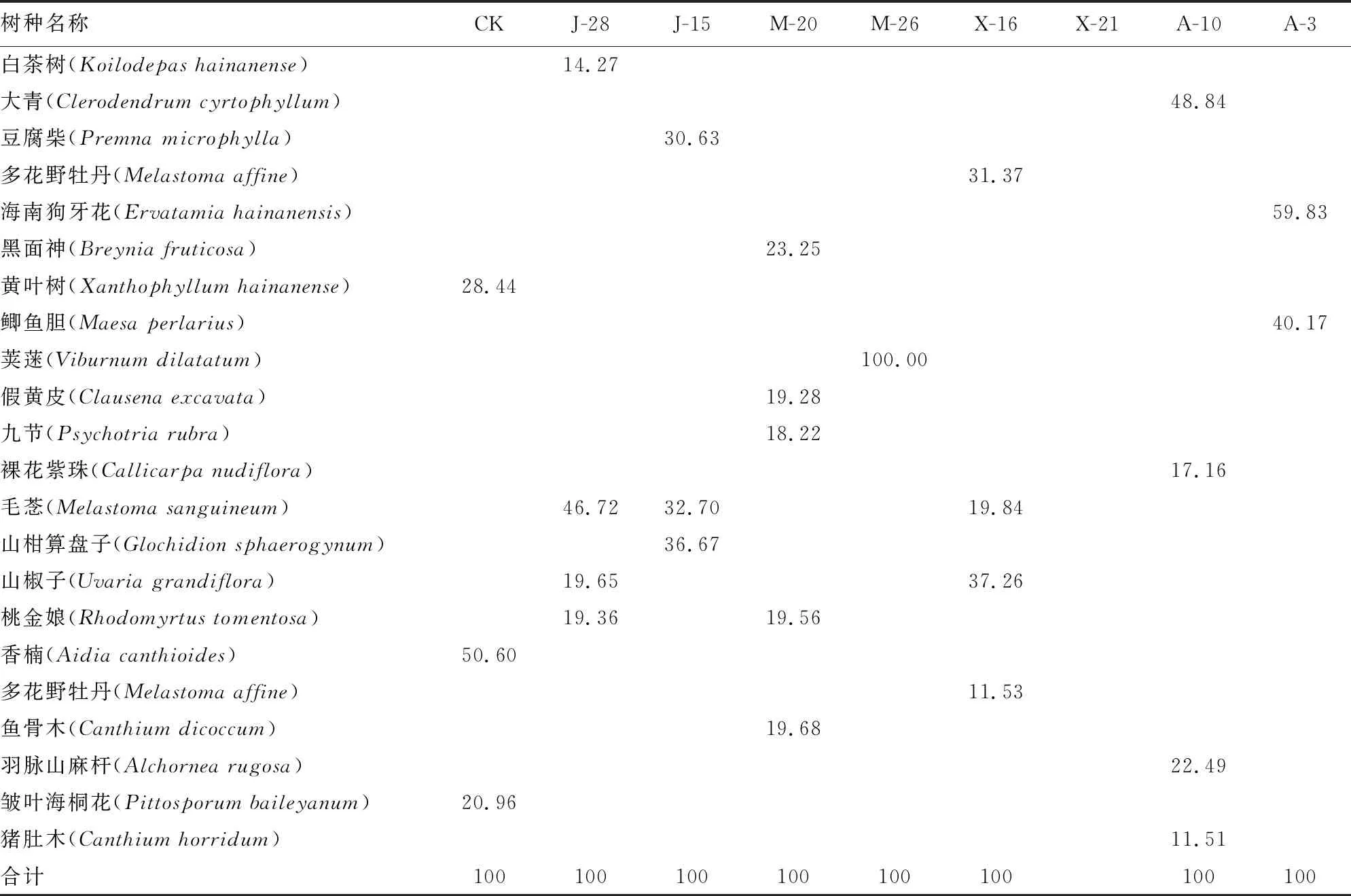
表4 调查样地群落灌木层植物重要值
2.2.3 群落草本层植物重要值 由表2、表5可知,CK草本层植物为0;J-28、J-15、M-20、M-26、X-16、X-21、A-10、A-3草本层植物都较少,只有A-3草本层植物7种和X-16草本层植物3种,其余森林群落草本层植物种类只有2种或1种。在多种草本植物中,乌毛厥(Blechnumorientale)分别生长在多个群落之中,且其重要值最大,其中J-28群落草本层乌毛厥重要值为76.42%,J-15、M-20、X-16、X-21群落草本层乌毛厥重要值分别为35.17%、59.80%、41.90%和47.44%。桉树人工林没有发现乌毛厥生长,推测是因为桉树林下干燥,不适应乌毛厥生长。该群落草本层植物相对稀少,且植物分布不均匀,不利于物种丰富度提高。
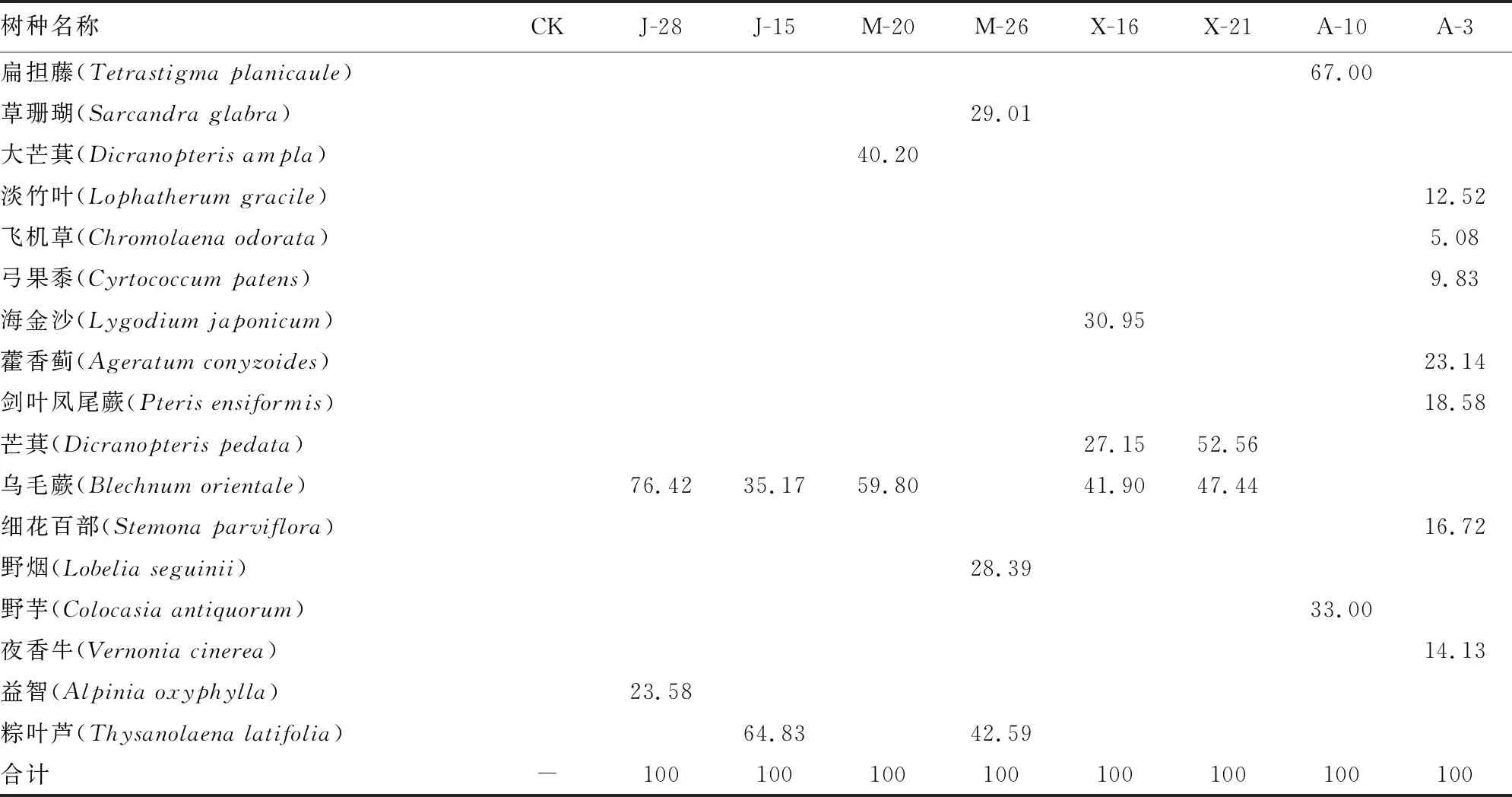
表5 调查样地群落草本层植物重要值
2.3 群落物种多样性分析
由表6可知,黎母山X-21群落乔木层、灌木层和草本层物种单一,同时A-3群落乔木层尚未形成,没法计算群落优势度指数(H)、多样性指数(H′)、均匀度指数(Jsw)。黎母山次生林群落与外来树种人工林群落物种H相差较大,总体各森林群落乔木层物种H大于灌木层和草本层;除X-21和A-3外,其余群落物种H呈现乔木层>灌木层>草本层;A-3群落物种H呈现草本层>灌木层。CK群落乔木层H最大,为0.92,其次为M-20群落乔木层H为0.88,表明CK群落乔木层物种复杂程度高于M-20群落乔木层。在各个群落灌木层H中,M-20群落灌木层H最大为0.80;草本层H显示A-3群落草本层H最大为0.81,各个群落草本层H相差不大。
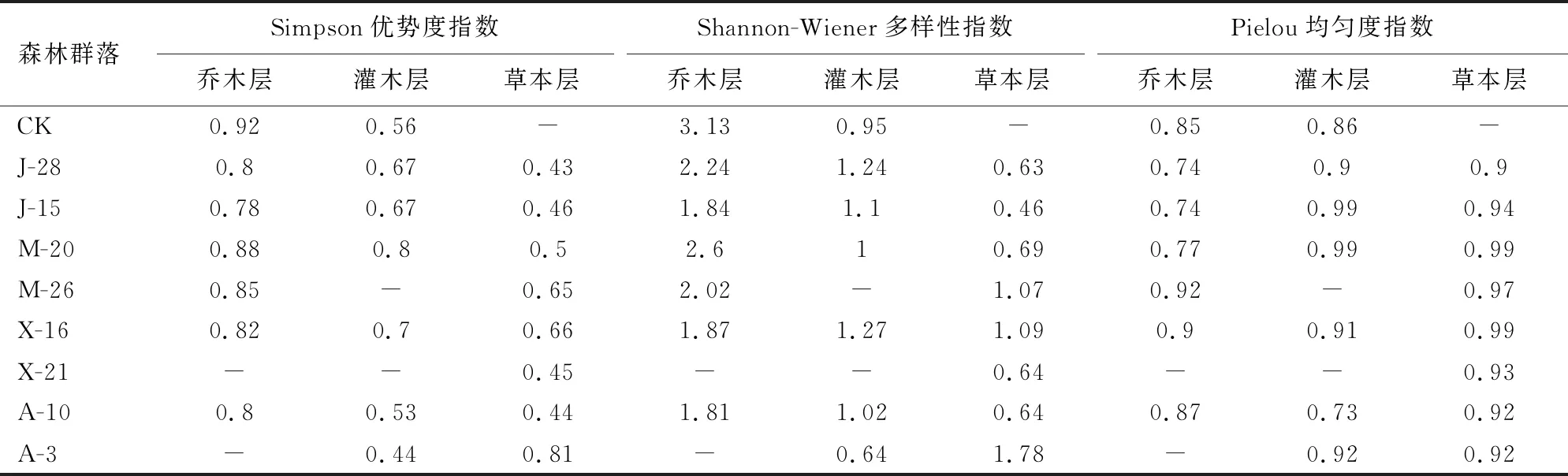
表6 森林群落不同层次物种多样性
黎母山次生林群落和外来树种人工林群落物种H′相差较大,除X-21和A-3之外,各个群落物种H′表现为乔木层>灌木层>草本层,A-3H′为乔木层>草本层>灌木层。群落乔木层H′最大为CK,H′为3.13。群落灌木层H′最大是J-28群落,其次是J-15群落,分别为1.24和1.10。草本层H′中,A-3群落最大,为1.78。
草本层和灌木层Jsw大于乔木层;部分群落的草本层和灌木层Jsw相差不大,其中J-28、A-3灌木层和草本层Jsw相等,分别为0.90、0.92,说明灌木层和草本层分布均匀情况接近,表明该群落草本层植被灌木层分布均匀程度大于草本层。群落乔木层物种Jsw最大是M-26群落,Jsw为0.92;灌木层物种Jsw最大是M-20,为0.97;草本层物种Jsw最大是X-16,为0.99。Jsw越大说明植物分布越均匀,反之越复杂。乔木层均匀指数小于灌木层和草本层,则反映乔木层复杂程度高于灌木层和草本层。
2.4 调查样地森林群落更新层树种
群落幼树种类调查结果见表7。次生林群落更新层乔木树种幼树种数为9个;J-28和J-15群落更新层乔木树种幼树种数分别为8个和7个;M-20和M-26群落更新层乔木树种幼树种数分别为21个和9个;X-16和X-21群落更新层乔木树种幼树种数分别5个和12个;A-10和A-3群落更新层乔木树种幼树种数分别为2个和3个。森林群落更新层树种调查结果没有发现对应人工林建群树种幼树,说明4个外来树种林分自然更新能力差,原因是林分郁闭度大,光照不足或树种自身繁殖方式。调查发现,在加勒比松和马占相思林群落出现海南木莲、白木香、乌檀、坡垒和红花天料木生长,在X-21群落出现乌檀生长,在次生林群里有白木香、青梅等珍贵树种生长。珍贵树种生长在树龄较大林分,表明该林分开始新的演替。外来树种人工林建群没有人为干预情况下,如果种群得不到更新,必将被林下乔木演替取代为混交林。海南珍贵树种以大径材为经营目标,生长速度慢,通常为耐阴树种,海南木莲、白木香、乌檀、坡垒和母生在树龄较大的林分中生长,则群落已经满足珍贵树种生长条件,若人为干预,随着珍贵树种生长繁殖,在将来生长为顶级群落优势群,完成珍贵树种自然回归过程。

表7 群落乔木幼苗种类
3 结论与讨论
3.1 结论
1)植被物种数量与组成具有差异。次生林乔木层物种数为40个,是调查到物种数最多群落;21 a橡胶群落和3 a桉树群落没有调查到乔木层树种。
2)森林群落乔木层优势树种具有差异。次生林分优势树种有贡甲和青梅;28 a加勒比松群落乔木层优势树种为鸭脚木和水锦树;15 a加勒比松群落乔木层优势树种为中平树、水同木;20 a马占相思群落乔木层优势树种为细齿叶柃;26 a马占相思群落乔木层优势树种为八角枫,16 a橡胶林群落乔木层优势树种是白楸,10 a桉树林群落乔木层优势树种为细基丸。
3)森林群落植被物种多样性具有差异。次生林物种多样性Simpson优势度指数为0.92,Shannon-Wiener多样性指数为3.13,Pielou均匀度指数为0.85,其群落物种多样性复杂程度高于外来树种人工林群落。20 a马占相思群落Simpson优势度指数为0.88,Shannon-Wiener多样性指数为2.60,Pielou均匀度指数为0.77,外来树种群落树种复杂程度最高,但不及次生林。由群落多样性指数反映,次生林乔木层物种多样性复杂,外来树种群里乔木植物组成复杂程度不及次生林群里,但灌木层和草本层相差不明显,表明次生林群落演替程度高于外来树种群落。
4)4个外来树种群落没有该树种幼树,更新层出现海南珍贵树种幼树,表明该建群树种的自然更新能力弱,随着群落的演替,现有的建群种可能被新的植物替代。因此通过人工修复外来树种群落,保持其群落应有的生态和经济价值是十分有必要的。
3.2 讨论
引入外来树种对一定区域内生态系统不一定是有害的[1],相对乡土树种,外来树种生长速度快,能在短时间内郁闭成林,对林地水分和养分的保持有着积极的作用[16-17],短时间内促进土壤养分的循环[9]。我国引入外来树种主要栽培区在热带和亚热带地区[18],以桉树、相思等速生树种为重要引入对象,同时也是海南省重要用材树种。相思类以马占相思最为活跃,马占相思具有良好的用材,同时马占相思具有根瘤菌,能固氮保湿[19],提高土壤微生物多样性[20],保持土壤养分[21]。黎母山引入马占相思后,乔木层物种多样性提高,马占相思林群落内出现海南珍贵树种红花天料木(母生)、坡垒和花梨木,说明马占相思引入特定区域对该乡土树种的影响是有利的,该现象与刘敏等[22]、廖宇杰等[23]结果一致与张宁南等[24]的研究结果有差异,原因推测是地理位置的差异造成。根据调查结果显示,28 a加勒比松群落物种丰富度与次生林群落乔木层、灌木层和草本层没有差异,但15 a加勒比松林群落较次生林群落乔木层物种丰富度较差,该结果总体与张快富等[25]的研究相似。
外来树种营造往往是人工纯林,而有学者认为引种外来树种形成人工纯林耗水量大、水分消耗快[4],不利于物种丰富度的提高[26],也有认为引种桉树对物种多样性的提高不如土著树种物种多样性提高效果优异[27]。但在调查结果中发现,加勒比松林群落和马占相思林群落中出现海南木莲、坡垒、乌檀、土沉香(白木香)和母生(红花天料木)生长,且马占相思群落植被多样性是高于次生林的,该现象说明外来树种成长到一段时期后,植被种类会发生更新,植被以乡土树种开始新的演替阶段,若无人为干预,珍贵乡土树种成为顶级演替群落优势树。若珍贵树种在自然条件中成林成材,所需要的时间比较长,需要人为干预,完成自然回归过程。
