水稻粒长新基因GL12-1定位与利用分析
2022-10-01何旎清黄凤凰余敏祥杨德卫
何旎清,黄凤凰,余敏祥,杨德卫
(福建省农业科学院 水稻研究所,福州 350018)
水稻(OryzasativaL.)为人类最重要的粮食作物之一,全球超过一半以上的人口以稻米为主食。据预测,到2030年世界人口将达到80亿,届时全球对水稻产量的需求将比现在增加约40%[1]。随着人民生活水平的提高,水稻粒型已成为当前消费者与育种家共同关注的热点,提高产量和改良品质已成为现代水稻育种两个主要目标[2]。
水稻粒长是水稻粒型构成因素之一,并直接影响水稻的单产,其中粒长对水稻粒重贡献最大[3-4]。有研究表明,长粒水稻品种米质相对较好[5]。随着人们生活条件的改善,优质稻米越来越受到人们的喜爱,研究表明,长粒稻米通常表现较好的外观品质[6]。粒长是稻米品质重要指标之一,水稻粒长相关研究已成为当前水稻遗传学家和育种家研究的热点。
据不完全统计,目前鉴定与粒长相关的QTL有120多个(http://www.gramene.org/)。由于粒长性状受环境影响大,研究者利用不同的研究方法与材料,鉴定的QTLs之间会存在重复性[7-8]。目前已有PGL1、GS2和qTGW3等[9]12个粒长相关基因被克隆。研究发现qTGW3有2个等位基因TGW3和GL3.3,qTGW3编码一个类GSK3/SHAGGY家族成员的激酶OsSK41/OsGSK5,能与生长素应答因子OsARF4互作并将其磷酸化[10]。TGW3编码一个类似糖原合酶激酶GSK3的激酶,是粒长的负调控因子,能协同改变颖壳中细胞的大小和数量[11]。GL3.3与之前发现的GS3在遗传上有上位互作效应,两者叠加能导致水稻粒型显著增大[12]。
目前虽然克隆了一些水稻粒长相关的基因,但有些基因存在与不良性状连锁的现象,从而限制这些基因在生产中的应用范围[13]。研究显示,APG是粒长基因PGL1拮抗性互作因子,APG是一个负向调节子,其功能受PGL1的抑制[14]。因此,鉴定和分离更多水稻粒长基因对水稻品质遗传改良以及揭示粒形性状遗传调控机理具有极其重要的意义。
本研究利用前期创制的长粒恢复系材料R20-4,通过正向遗传学的方法,对该长粒性状进行基因鉴定与定位分析,同时将恢复系R20-4分别与三系不育系‘庆源A’、‘定源A’、‘启源A’和‘靓香A’进行测交,并对测交组合的粒长进行分析。本研究可为后期GL12-1基因的克隆、功能研究以及育种利用研究提供参考。
1 材料和方法
1.1 材料及种植
长粒恢复系R20-4为含有稻瘟病抗性基因Pigm-1的恢复系,由福建省农业科学院水稻研究所通过分子标记辅助选择选育而成[15]。CO39为福建省农业科学院水稻研究所种质创制研究室保存的遗传材料。‘庆源A’、‘定源A’、‘启源A’和‘靓香A’为福建省农业科学院水稻研究所选育的三系不育系。
2019年8月R20-4与CO39进行正反交,2019年12月进行海南加代繁殖,2020年7月R20-4、CO39以及构建的F2群体均种植在福建省农业科学院水稻研究所连坂试验农场。两个亲本分别种植1个小区,每个小区设置3个重复,每个重复种植4行,每行8株,所有材料均单株种植,按照常规的栽培技术进行管理。
1.2 方法
1.2.1 长粒性状的遗传分析将R20-4与CO39进行正反交,待群体完全成熟后,根据F1的粒型表型,以及F2代分离群体中长粒单株与短粒单株的调查比例,来分析长粒性状的遗传特性。
1.2.2 亲本与F2性状调查分析对R20-4和CO39粒长性状进行调查分析,稻谷粒长等测定参照Tan等[16]方法进行。成熟后,对正交F2群体中极端长粒单株和短粒单株进行取样,选择极端长粒20株和极端短粒20株作为连锁群体,正交F2群体中获得的短粒单株作为后期的定位群体。
1.2.3 长粒基因初步定位水稻组织DNA的提取、PCR的扩增以及样品电泳检测参照杨德卫等[17]方法。利用实验室506对SSR引物对R20-4和CO39进行多态性分析,利用筛选的多态性引物,对粒长基因进行连锁分析。
1.2.4 长粒基因精细定位标记的开发和对应分子标记的物理距离均参照杨德卫等[17]方法。利用粳稻品种‘日本晴’和籼稻品种‘9311’两品系已公布的核苷酸序列(http:// www.ncbi.nlm.nih.gov),找到序列之间有差异的部分,差异大小一般选择在50 bp以下,然后利用引物设计软件Primer 5.0在其两侧设计引物。
1.2.5 长粒基因育种利用研究将恢复系R20-4分别与‘庆源A’、‘定源A’、‘启源A’和‘靓香A’三系不育系杂交,并对配制组合F1的粒长性状进行调查分析。
2 结果与分析
2.1 亲本粒长性状比较
为选择定位群体的亲本,本研究对恢复系R20-4和CO39粒长进行调查分析显示,恢复系R20-4粒长为(11.31±0.25) mm,CO39粒长为(8.25±0.18)mm(图1),分析表明R20-4的粒长与CO39相比,差异达到极显著水平 (P<0.01)。
2.2 恢复系R20-4长粒性状的遗传分析
为了分析R20-4长粒性状的遗传特性,本研究将R20-4和CO39进行正反交。结果显示,F1植株均表现长粒的表型。将正反交F1自交收获后种植,成熟后调查F2群体粒长性状分离情况,长粒与短粒单株的分离比经卡平方(χ2)检验符合孟德尔遗传3∶1比例(表1),粒长性状在正反交F2群体中呈现双峰分布(图2)。以上结果说明R20-4长粒性状是由1对显性基因控制的。
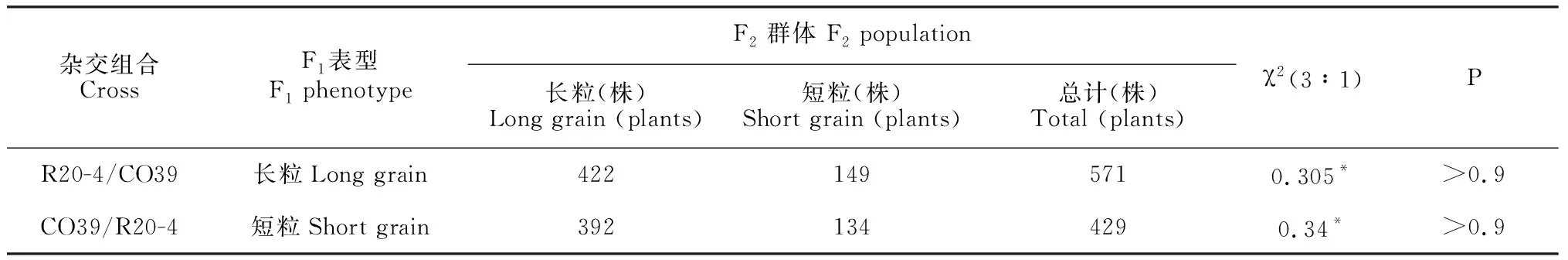
表1 长粒性状的遗传分析
2.3 R20-4长粒基因的初步定位
为了定位恢复系R20-4长粒基因所在染色体的位置,本研究利用实验室合成的506对SSR和Indel引物标记,对R20-4和CO39进行多态性筛选,有96对引物在两个亲本之间存在多态性。利用这96对引物对F2群体中的20个极端长粒单株和20个极端短粒单株进行连锁分析。发现水稻第12染色体长臂上标记Indel-12-3和Indel-12-7可能与粒长基因存在连锁。对F2群体中45个小粒单株进一步分析,发现Indel-12-3和Indel-12-7与短粒位点存在连锁关系。最终利用314个短粒隐性单株,将该长粒基因位点定位于水稻第12染色体上标记Indel-12-3和Indel-12-7之间,物理距离约4.5 Mb,并命名为GL12-1(图3,A)。
2.4 长粒基因进一步定位
本研究在Indel-12-3和Indel-12-7之间合成20对Indel引物,多态性检测显示只有4对引物具有多态性(表2),分别为Indel-3、Indel-6、Indel-10和Indel-16,利用这4对多态性引物对314个短粒隐形单株进行分析,最终将GL12-1基因定位在标记Indel-10和Indel-16之间,物理距离约为900 kb (图3,B)。
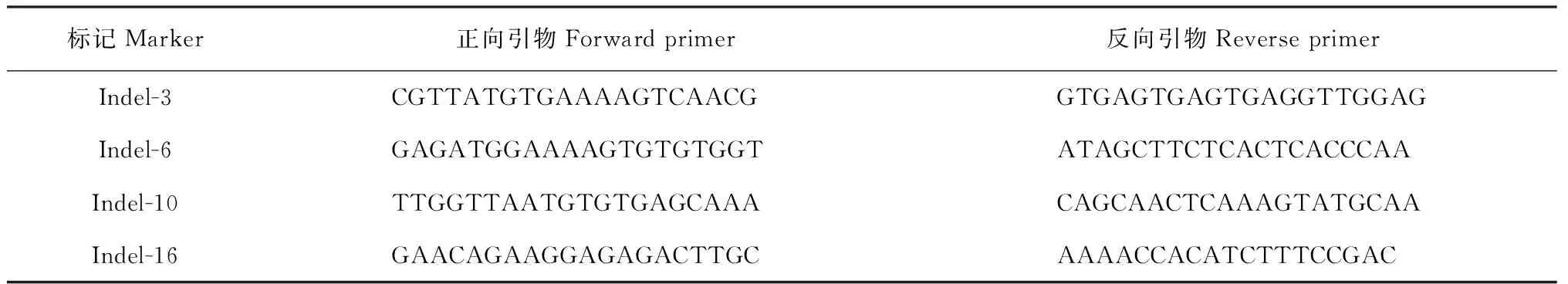
表2 Indel引物
2.5 R20-4长粒基因的利用
2021年7月分别将R20-4以及配制的4个杂交组合庆源A/R20-4、定源A/R20-4、启源A/R20-4和靓香A/R20-4种植在福建省农业科学院水稻研究所连坂试验农场,待成熟后对其粒长性状进行分析,并对其对应的不育系粒长性状进行分析。结果表明,组合庆源A/R20-4、定源A/R20-4、启源A/R20-4的粒长与R20-4相比,没有明显差异,但靓香A/R20-4的粒长与R20-4相比,表现粒长变长,差异达到显著水平(表3)。
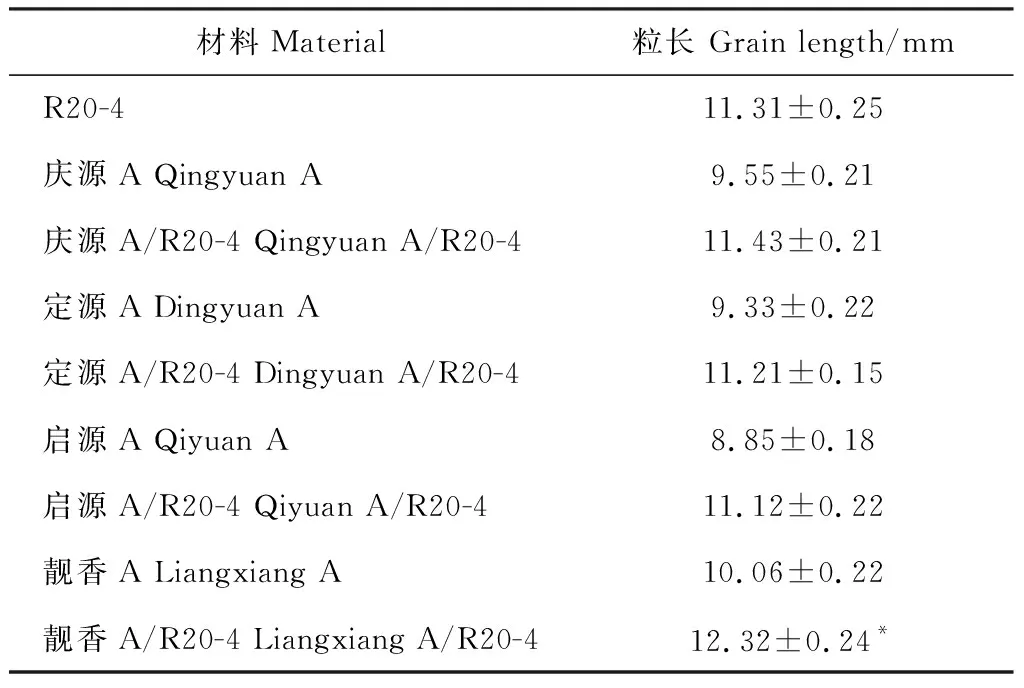
表3 恢复系R20-4及其配制组合的粒长分析
3 讨 论
3.1 GL12-1可能是一个新的粒长控制基因
目前为止,已发现120多个与粒长相关的基因或QTL (http://www.gramene.org/)分布于水稻不同染色体上,主要位于第1、2、3、7和10号染色体上。本研究利用图位克隆的方法,将R20-4长粒主效基因GL12-1进行了遗传定位,进一步分析发现该区间还没有克隆的粒长相关基因。因此,我们推测GL12-1可能是一个新的控制粒长相关基因。
GL12-1候选基因区间900 kb内包含有100多个候选基因,所以目前还无法确定哪个基因为其候选基因。另外,由于CO39与R20-4均为籼稻材料,他们之间的遗传差异较小,多态性引物少。本研究在进一步定位过程中,合成的20对引物仅有4对具有多态性。随后,我们在Indel-10和Indel-16标记之间又合成8对Indel引物,检测结果显示这8对引物均没有多态性。目前我们正在构建遗传差异较大群体,以期完成GL12-1基因的精细定位及克隆工作。
3.2 GL12-1基因可能具有较好的育种利用价值
水稻粒长性状在不同水稻品种之间存在多样性变化,一般在6~15 mm之间。例如粳稻品种‘川7’的粒长仅有7.32 mm,而籼稻代表品种‘明恢63’粒长达到10.2 mm[9]。水稻粒长性状,由于材料遗传背景的影响,其遗传特性也不尽相同,主要受多基因控制以及由主效基因加微效基因共同调控[18-19]。
在遗传学方面,粒长的遗传基础极为复杂。研究者利用3个长粒材料与3个短粒材料配制不同的杂交组合,研究表明粒长性状为多基因控制的数量性状,其加性效应占比较大[20];周清元等[21]研究表明,杂种F1的粒长介于两个亲本之间;贾小丽等[22]利用重组自交系群体(recombinant inbred lines,RILs) 研究发现,粒长性状呈连续分布,且表现出明显超亲分离现象。
在基因功能方面,粒长基因调控遗传机理极为复杂。有的为正调控,如GLW7[23];有的为负调控,如GS3[24];有的由于氨基酸变化,导致粒长变化,如qGL3[25];有的由于结构变异,导致功能丧失,从而引起粒长变化,如qTGW3[11];有的由于基因转录提前终止,导致产生不同大小蛋白,从而引起其粒长变化,如GL3.2[26];有的与某些蛋白或者转录因子互作,引起蛋白表达变化,进而影响粒长变化,如Gnp4[27-28]。
本研究利用长粒材料R20-4,在配制的4个不同不育系杂交组合中,有3个组合表现亲本R20-4粒长特性,有1个组合表现超亲遗传的特点,推测长粒基因GL12-1以显性遗传为主。配制的杂交组合R20-4/靓香A表现超亲遗传现象,原因可能是在R20-4与靓香A形成杂交组合中,由于有些基因编码蛋白之间存在互作,或者微效基因叠加,从而影响或调控其组合粒长变化。
因此,本研究鉴定的长粒基因GL12-1在今后粒型育种可能具有较广的应用价值。尤其在杂交水稻选育过程中,两个亲本只需一个亲本含有GL12-1,其组合就表现长粒表型。后期我们将通过对GL12-1基因进一步克隆,开发功能性分子标记,并结合常规杂交技术,以期快速有效地聚合多个基因的育种目标,这不仅降低了育种成本,同时缩短了育种时间。
最后,随着国家的快速发展和社会的进步,人们已经步入小康社会,在吃饱的同时人们开始追求吃好、吃健康。长粒水稻品种在外观及口感等方面受到人们的喜爱[10, 13, 29]。一方面,在粒型形成遗传学方面取得很大进展,对粒型性状遗传及基因功能方面认识较为明晰[10, 29]。而另一方面,对水稻粒型基因调控机理方面的研究相对较少,如在大面积田间推广长粒品种并达到产量提升的效果更是少之又少。因此,研究者需进一步加大水稻粒长种质创新、遗传规律探索以及调控机理等基础方面研究,这对于加速水稻粒长新品种选育以及解决粮食问题具有重要意义。
本研究通过图位克隆的方法,对水稻长粒基因GL12-1进行了鉴定,该基因可能是一个新的粒长基因。与不育系测交表明,GL12-1具有显性遗传特性,该基因在今后的水稻粒型育种中可能具有较广泛的应用价值。
