不同居群红砂的遗传多样性分析
2022-09-29苏世平师微柠AlizetDidiDom
张 旭, 苏世平, 师微柠, Alizet Didi Dom, 马 强
(甘肃农业大学, 甘肃 兰州 730070)
红砂(Reaumuriasoongorica)隶属于柽柳科(Tamari-caceae)石竹亚纲(Caryophyllidae)[1],主要分布于亚欧大陆(中亚)、南欧和北非,共有21种,亚洲有18种,我国分布有4种,多产自内蒙古和西北地区[2]。红砂是我国干旱荒漠区分布最广的植物种之一,也是荒漠化、半荒漠化地区防风固沙和水土保持的优良灌木树种,和荒漠绿洲与荒漠过渡带的重要植被类型,其具有抗旱、耐旱、耐盐碱及很强的集沙固沙能力[3-4]。
近年来,由于干旱和高温等极端气候的频发,导致红砂种子繁殖困难,长期的无性繁殖导致遗传多样性降低[5],因此探讨红砂种群的遗传变异规律尤为重要,为其遗传资源的保护和评价提供理论依据。李秀玲等[6]运用空间自相关对我国西北地区的28个红砂种群进行空间分布的研究,结果表明红砂种群的空间分布模式为随机分布。徐莉等[7]应用了RAPD分子标记,15对引物对分布于新疆阜康荒漠7个亚群的136株红砂进行扩增。Shannon多样性指数(0.307)和居群间遗传分化系数(0.312)揭示了红砂的遗传变异多存在于亚种群内,亚种群间的遗传分化较小。殷恒霞等[8]采用磁珠富集法构建了红砂基因组微卫星富集文库,分析得出微卫星重复基元显示为5种类型,其中三碱基基元(GGT)n,(TGG)n,(ACC)n最多,占总数的60.7%。杨九艳等[9]运用ISSR对内蒙古高原荒漠区红砂居群的遗传多样性进行研究,结果表明红砂居群间有高度的遗传分化。张颖娟等[10]应用RAPD分子标记对内蒙古西部红砂种群进行遗传多样性分析,结果表明不同生境下红砂种群内存在较高的遗传多样性,聚类分析表明遗传距离与地理距离之间存在一定相关性。
本研究基于前人的研究基础,应用SSR分子标记对来源于不同区域的16个红砂居群进行多样性水平评估及群体遗传结构分析,从分子水平上探究红砂居群的遗传规律,以期为红砂的合理利用及有效保护提供理论基础。尽管在红砂遗传多样性研究进展中有相关研究,但是都基于小地域,而对全国大地域分布区的研究报道较少[11-14]。应用的SSR分子标记具有高度重复性,丰富多态性及共显性的优点,为目前应用比较广泛的分子标记技术[15]。
1 材料与方法
1.1 试验材料
2011年11月在我国甘肃、宁夏、内蒙古的红砂天然分布区的各居群中,随机选取红砂植株并采集种子,标记、分装;2012年1月在武威市林业技术服务中心苗圃利用所采集的种子进行穴盘育苗;2013年4月土壤解冻后进行移栽,并按常规方法进行育苗期管理,建立了位于武威市凉州区下双镇国家樟子松基地向北1 000米的红砂试验地。本研究实验材料均取自该试验基地。
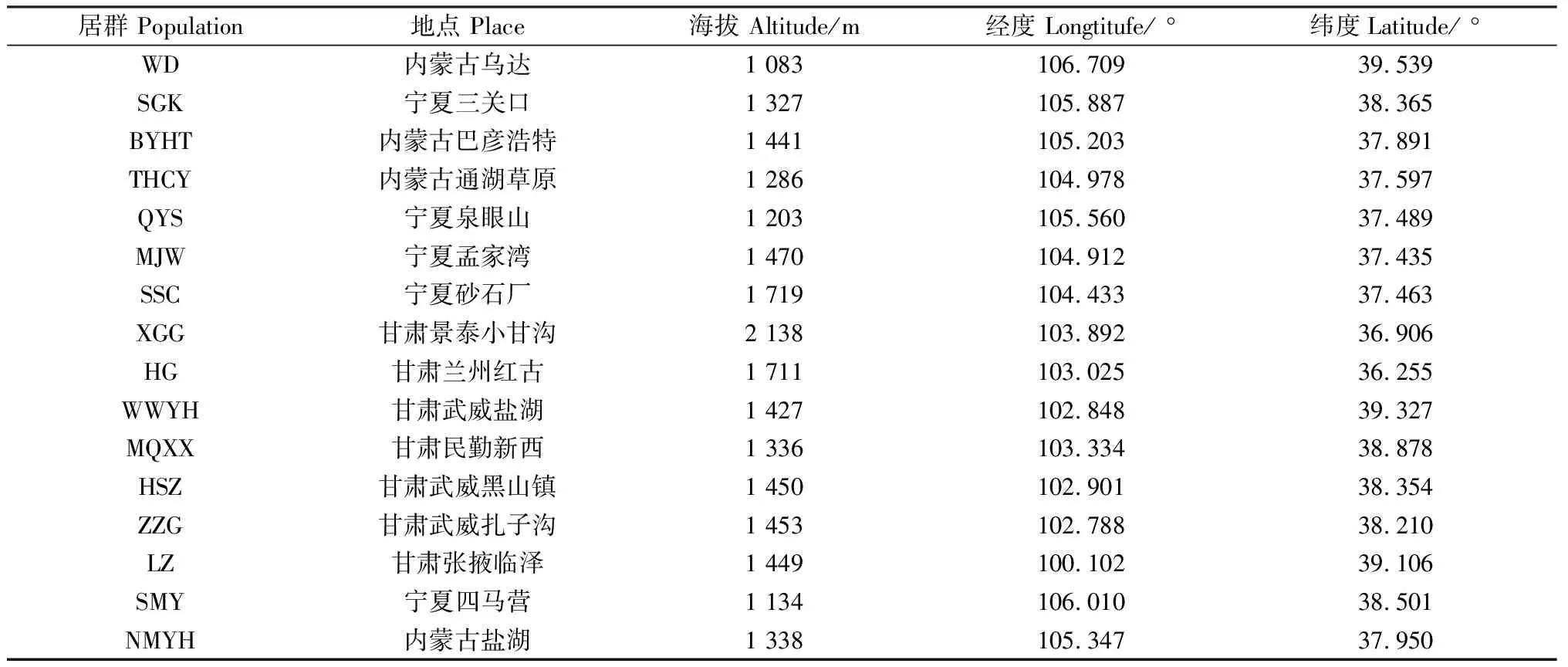
表1 供试材料红砂不同居群信息Table 1 Information of different populations of R. Soongorica
1.2 DNA提取及扩增
用磁珠法基因组提取试剂盒(天根生化科技有限公司)根据产品说明按步骤进行总DNA的提取。从96对引物中筛选出11对多态性较高的引物(SSR引物由武汉天一辉远生物科技有限公司合成)用于PCR扩增。反应体系为15 μL:2×Taq PCR Master Mix7.5 μL,Mix primer(引物混合液)2.0 μL,DNA template 1 μL,ddH2O 4.5 μL。反应程序:95℃预变性5 min;95℃变性30 s,52℃~62℃退火30 s,72℃延伸30 s,共10个循环;95℃变性30 s,52℃退火30 s,72℃延伸30 s,共22~25个循环;72℃延伸2 min;4℃保存[11-12]。
1.3 数据分析
通过群体遗传学软件GeneAlex6.41计算每个位点的等位基因数(Na)、有效等位基因数(Ne)、期望杂合度(Expected heterozygosity,He)、观察杂合度(Observed heterozygosity,Ho)、基因分化系数(Fst)、基因流(Gene Flow,Nm)、香农信息指数(I)等[16];并对群体间进行分子方差分析(AMOVA)[17-18]、居群间的遗传距离(Nei′s Genetic Distance,D)和遗传一致度(Nei′s Genetic Identity,GI)[19]。基于Nei′s遗传距离,用Phylip软件构建UPGMA聚类图[20]。
有效等位基因数为纯合度的倒数,是反应群体遗传变异大小的指标之一,其值越接近等位基因数的绝对值,表明等位基因在群体中分布越均匀[21]。式中,Pi为第i个位点上等位基因出现的频率。
Ne=1/1-He
香农信息指数用于估算居群内的遗传分化,指数越大居群间或居群内的分化程度越高,且遗传多样性越高[22]。
I=-∑pilnpi
期望杂合度(Expected heterozygosity,He),根据每个等位基因的基因频率计算,期望杂合度的值越高,表明物种多态性越高[23]。
多态信息含量指数用来衡量基因座多态性高低的程度[24]。式中,n代表的是样本中序列的数量。
自然群体并非是理想群体,如果仅考虑单基因座上一对等位基因,对于由若干个地方群体组成的生物大群体,各层次现实群体中基因型频率期望值偏差一般用Fst度量。计算公式如下[25]:
式中,群体内平均杂合度(The mean Heterozygosity within Populations,Hs),为各个居群的期望杂合度的算数平方根。
整体期望杂合度(Total Expected Heterozygosity,HT)的计算公式如下:
基因流代表等位基因从一个居群到另一个居群间的转移。基因流大于1,就能发挥均质化作用,能有效抑制有遗传漂变而引起的遗传分化[26]。
Nm=[(1/FST)-1]/4
2 结果与分析
2.1 位点多态性评估
11对引物扩增出67个等位基因,其中多态等位基因为56个,多态率为83.58%。扩增长度为126~396 bp,位点ReS078上扩增出的等位基因数最多,位点ReS008最少。多态含量指数为0.061~0.910,均值为0.396,位点ReS078的多态含量指数(0.910)为最高,位点ReS008的PIC指数(0.061)为最低。红砂基因位点的多态性水平为中度多态性(表2)。
2.2 居群多样性分析
所测红砂居群的等位基因数为2.364~4.455,均值为3.735,有效等位基因数为1.504~2.585,均值为2.221,结果显示居群的观测等位基因数均高于有效等位基因数(表3)。其中居群THCY的Na(4.455)和Ne(2.585)为最高,居群MQXX的Na(2.364)和Ne(1.504)为最低。观测杂合度为0.295~0.418,均值为0.358。居群LZ的Ho(0.418)为最高,表明该居群内杂合子占比为41.8%;MQXX的Ho(0.286)为最低,说明该居群内纯合子占比较多。居群观测杂合度和期望杂合度均小于0.5。仅有MQXX观测杂合度大于期望杂合度,在该居群内存在杂合度缺失的现象。香农信息指数为0.420~0.873,均值为0.763,说明居群内的遗传分化程度低,按照香农多样性指数评估红砂居群的遗传多样性水平:THCY,ZZG>LZ>NMYH>QYS>HSZ>WWYH>MJW>SSC>BYHT>HG>XGG>SGK>WD>SMY>MQXX。
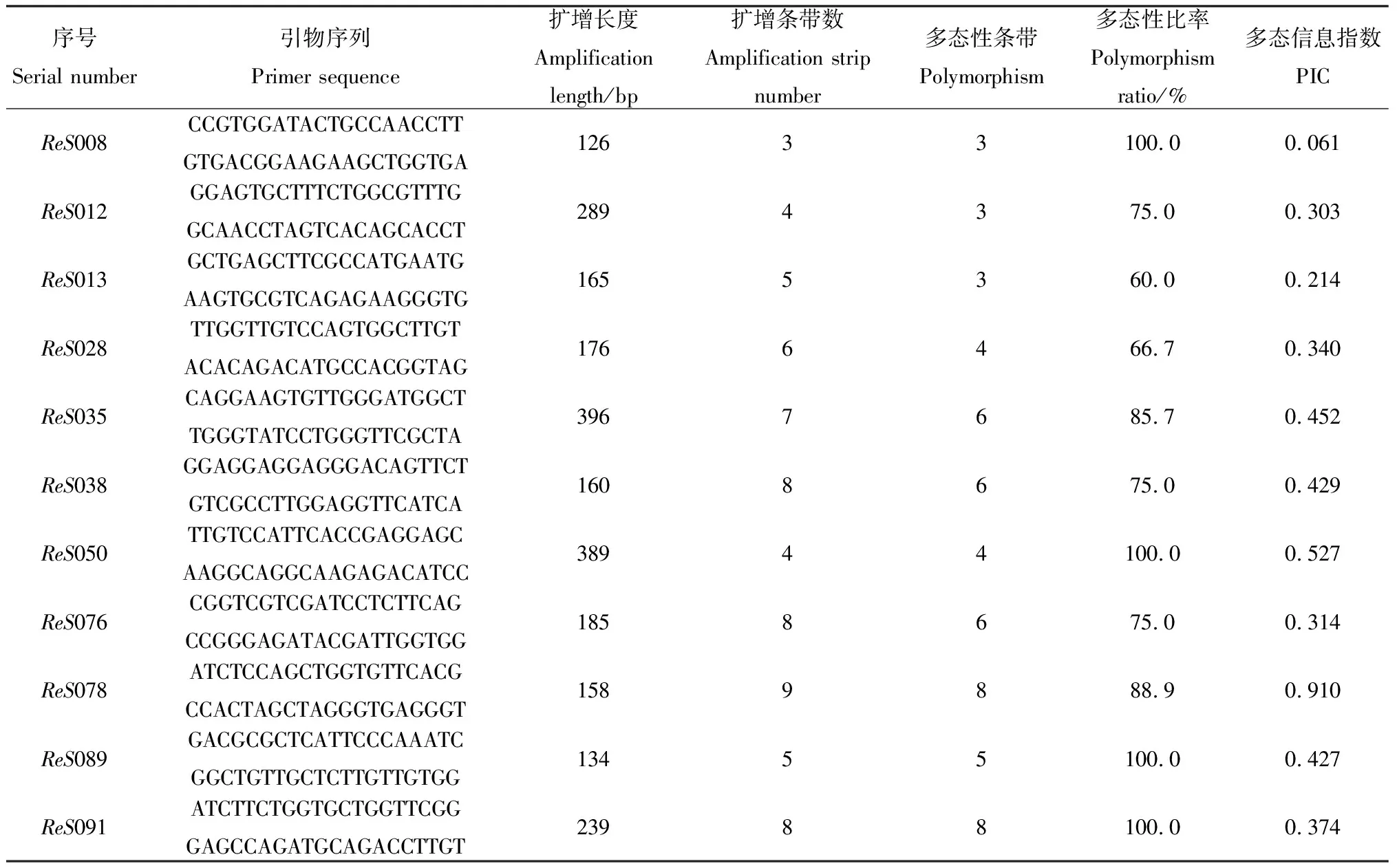
表2 11对引物多样性评估Table 2 Diversity Evaluation of 11 pairs of primers
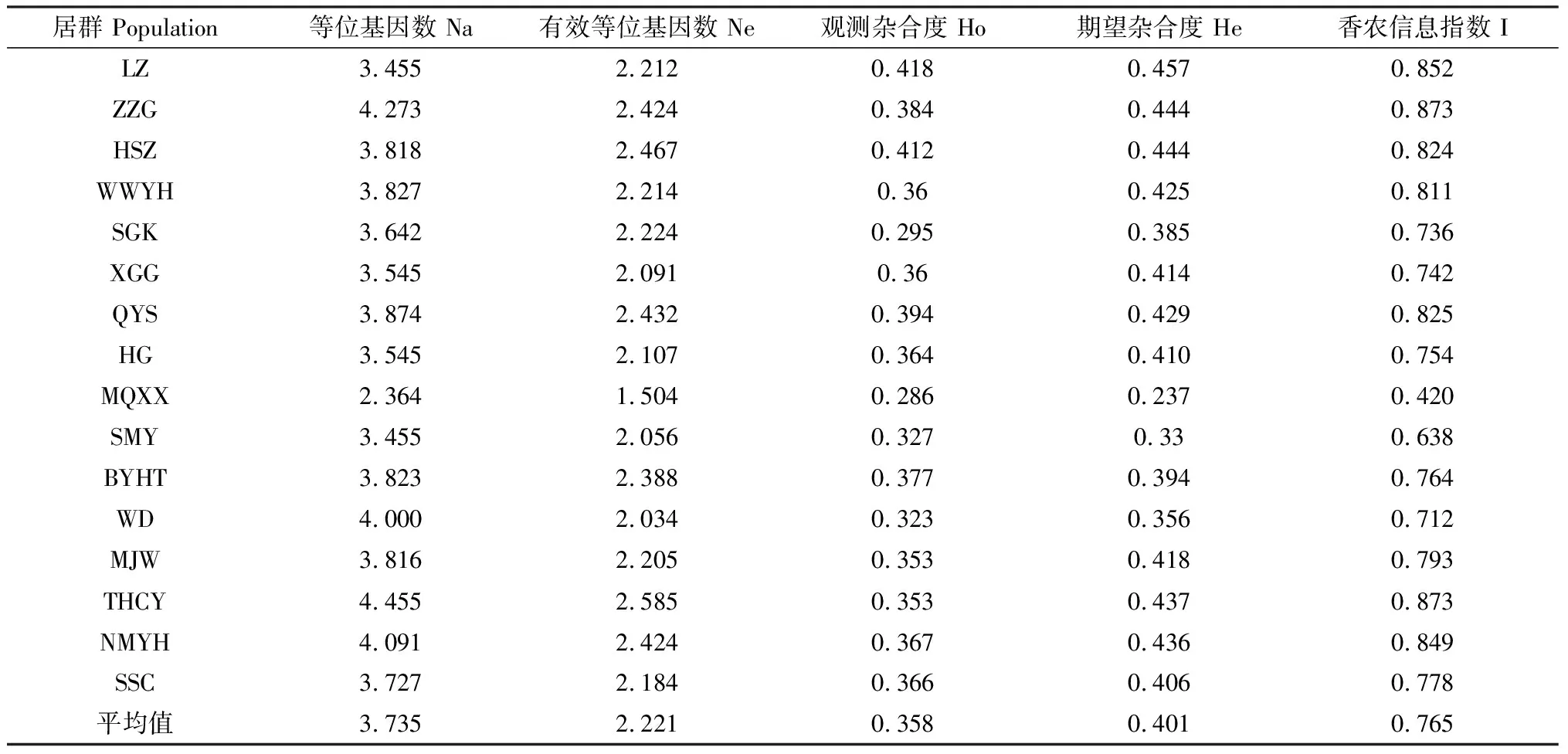
表3 居群各项多样性参数(均值)Table 3 Values of population diversity parameters(Mean value)
2.3 遗传分化及基因流
红砂整体近交系数为-0.026~0.548,整体表现为近亲交配,但在地方亚群内存在异交现象(表4)。居群间遗传分化系数为0.041~0.21,平均值为0.088,说明居群间遗传分化程度较低。AMOVA结果表明,居群间的遗传差异占变异总来源的5.8%,个体间占14.8%,个体内的遗传差异占79.4%,得出红砂的遗传变异材料主要来源于红砂个体内。各居群基因交流为0.940~5.807,平均值为3.163,该结果表明所测红砂居群间有较强的基因交流。
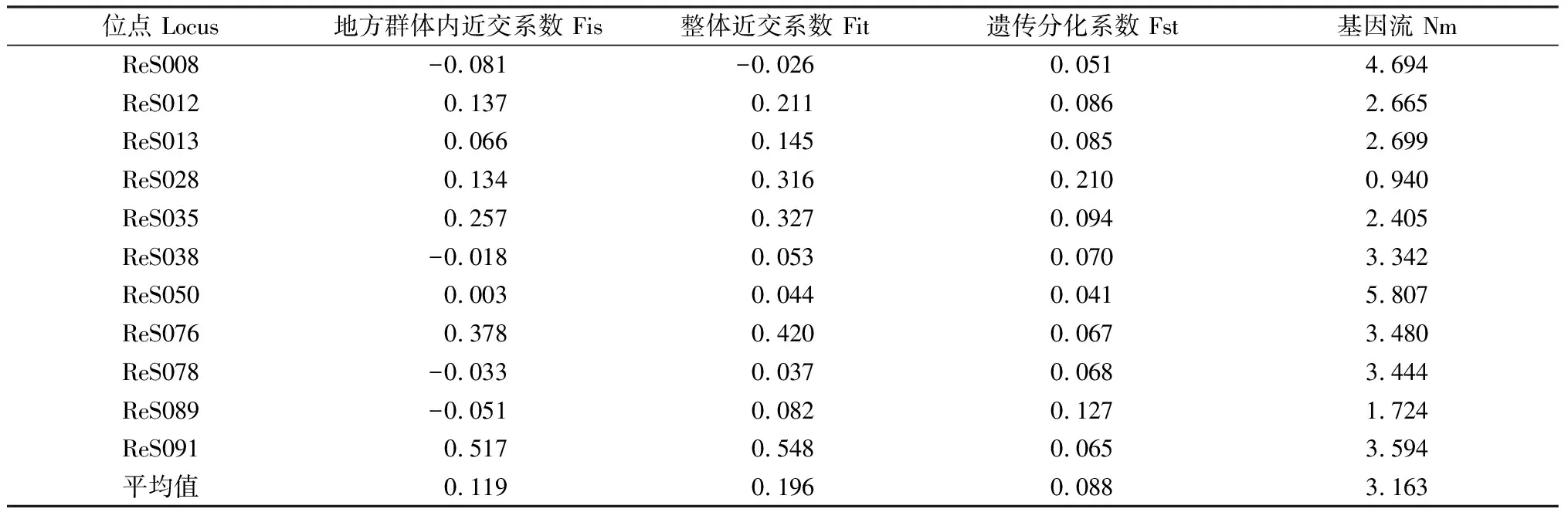
表4 各位点F-Statistics统计值和基因流Table 4 Each point F-statistics Statistics and gene flow
2.4 遗传结构及Mantel检测
红砂居群间的遗传距离为0.009~0.229,均值为0.062,遗传距离最近的居群是WD和SMY,最远的居群是MQXX和HG(表5)。遗传距离小于0.1的居群占总数的76.7%,大于0.1的占总数的23.3%,表明所测红砂居群间的遗传距离相近。居群间遗传一致度为0.795~0.983,均值为0.933,其中遗传相似性最大的居群是WD和SMY,最小的是MQXX和HG。Mantel检测(图1)得出红砂居群的遗传距离和地理距离显著性相关(R2=0.3254,P=0.02<0.05)。
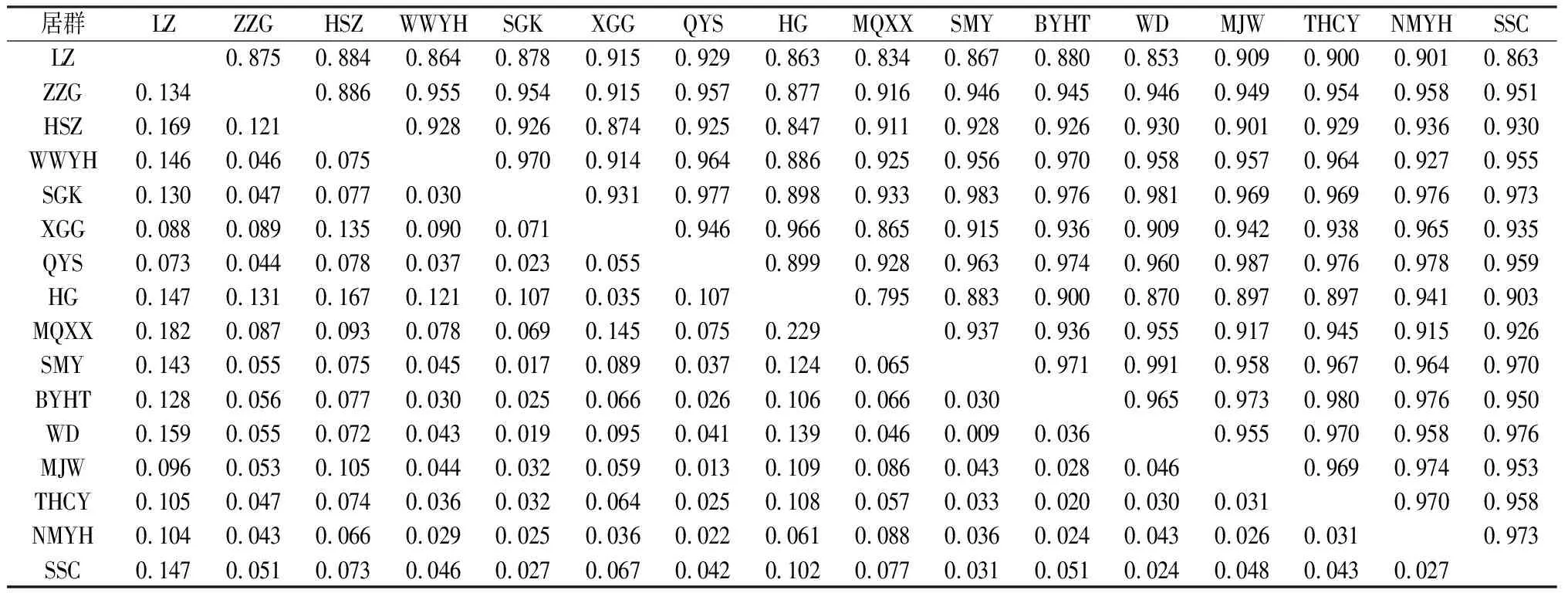
表5 遗传距离(左下)和遗传一致度(右上)Table 5 Enetic Distance (Lower left) and Genetic Identity (Upper right)
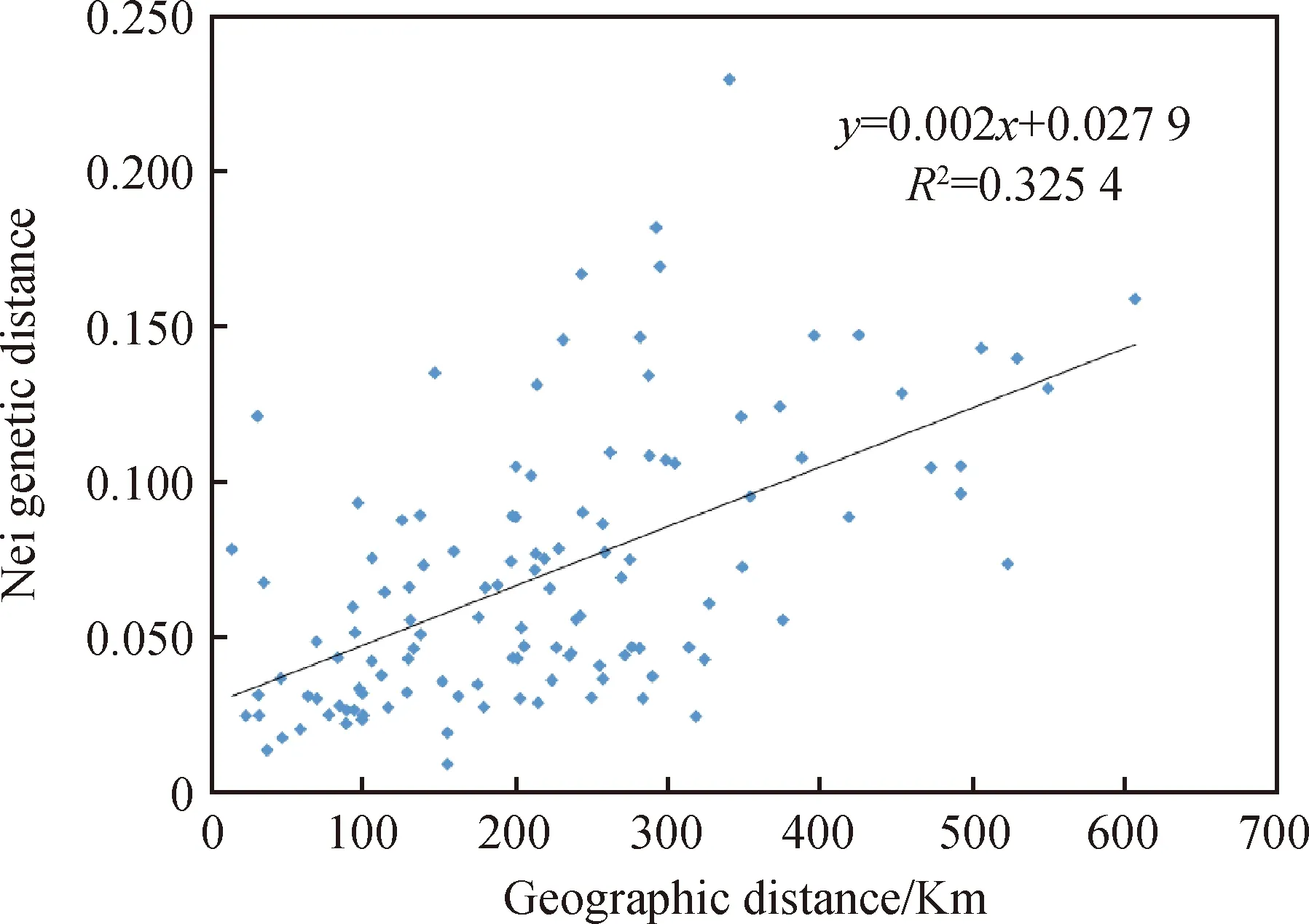
图1 遗传距离与地理距离的Mantel检测Fig.1 Mantel test of Genetic distance and geographical distance
2.5 聚类分析及PCoA主成分分析
基于红砂居群间的遗传距离并采用UPGMA进行聚类(图2)。SSC,SGK,WD,SMY,THCY,BYHT,NMYH,MJW,QYS,WWYH,ZZG被聚为一类;然后和MQXX聚成第二类;和HSZ聚为第三类;再和XGG,HG聚为第四类;最后和遗传距离较远的LZ聚为第五类。PCoA分析结果(图3)将红砂居群被分为5个部分。第一部分为HG,XGG;第二部分为NMYH;第三部分为LZ;第四部分为MQXX;其余居群为第五部分。PCoA与UPGMA聚类结果基本一致。
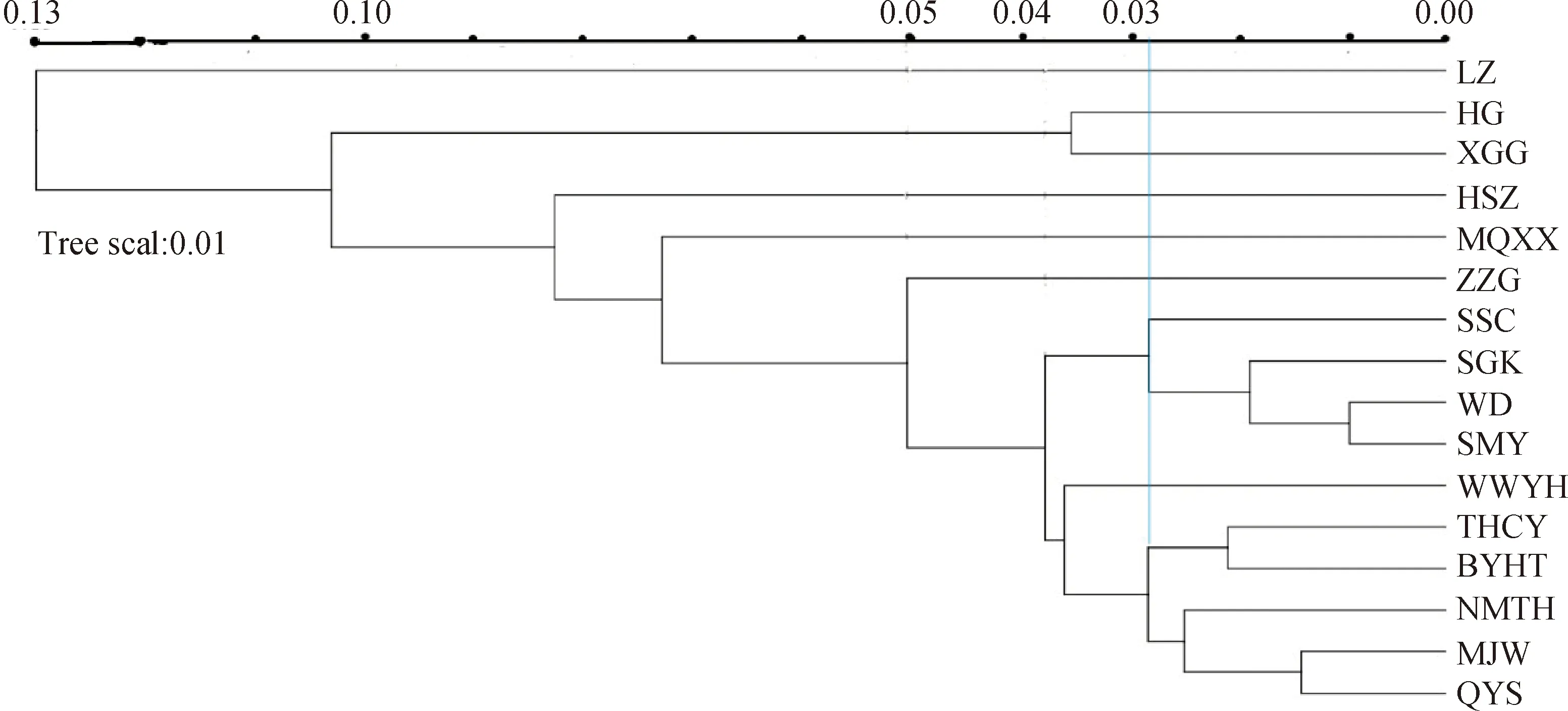
图2 不同居群红砂UPGMA聚类图Fig.2 UPGMA clustering diagram of R. soongorica in different cohabitation groups
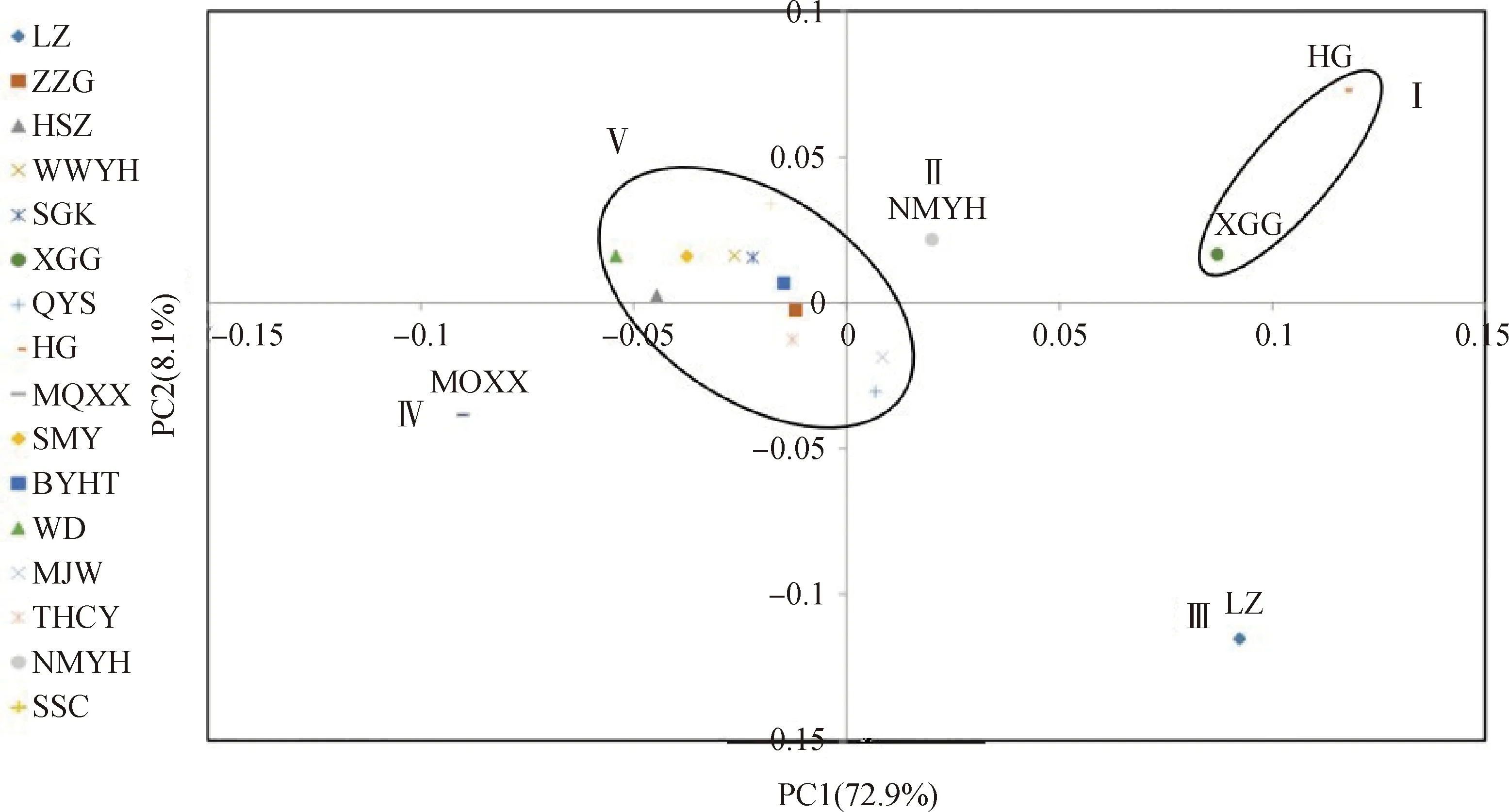
图3 不同居群的PCoA分析Fig.3 PCoA analysis of different populations
2.6 STRUCTURE结构分析
当K=2时,299份材料被分为2个亚群;当K=3时,能将MQXX,HSZ和XGG,HG区分开;模拟k值2~20,得出的最大似然值为5,说明299份红砂材料被分为5个亚群(图4)。第Ⅰ亚群(绿色)共75份材料,占总数的25%,第Ⅱ亚群(蓝色)共75份红砂,占总体的25%,第Ⅲ亚群(粉色)包含66份材料,占总数的22.1%,第Ⅳ亚群(橙色)包含材料56份,占比为18.7%。第Ⅴ亚群(黄色)共27份,占比为9%。聚类分析将299份材料分为5个亚群(图5),第Ⅰ类中LZ为4份,THCY,HSZ各3份,ZZG,MJW,SSC,QYS,SGK各2份。第Ⅱ类中与XGG,HG(占总数37.8%)分在一起的有46份材料,各占比为NMYH(9.2%),WWYH(7.8%),THCY(6.5%)。第Ⅲ类以MJW为主共计聚类23份材料。第Ⅳ类共有76份红砂材料,其中ZZG(13.1%),HSZ(15.7%),NMYH(9.2%),SSC(9.2%)聚在一起。第Ⅴ类共有106份红砂材料,占比为MQXX(15%),SMY(13.2%),WD(11.3%),BYHT和SGK(各为8.4%),THCY,SSC和QYS(各占6.6%),NMYH,ZZG(各占5.6%),MJW(4.7%),WWYH(2.8%),HSZ(1.8%),XGG(0.9%)。利用聚类分析校对了红砂的亚群分类,两者结果显示299份红砂材料被分为5个亚群。
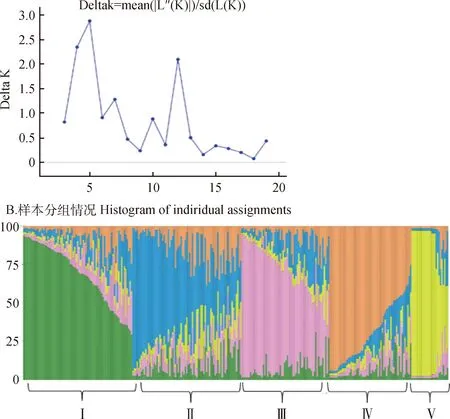
图4 不同k值的遗传结构与Δk值的折线图Fig.4 Genetic structure of different K values and the magnitude of Δk
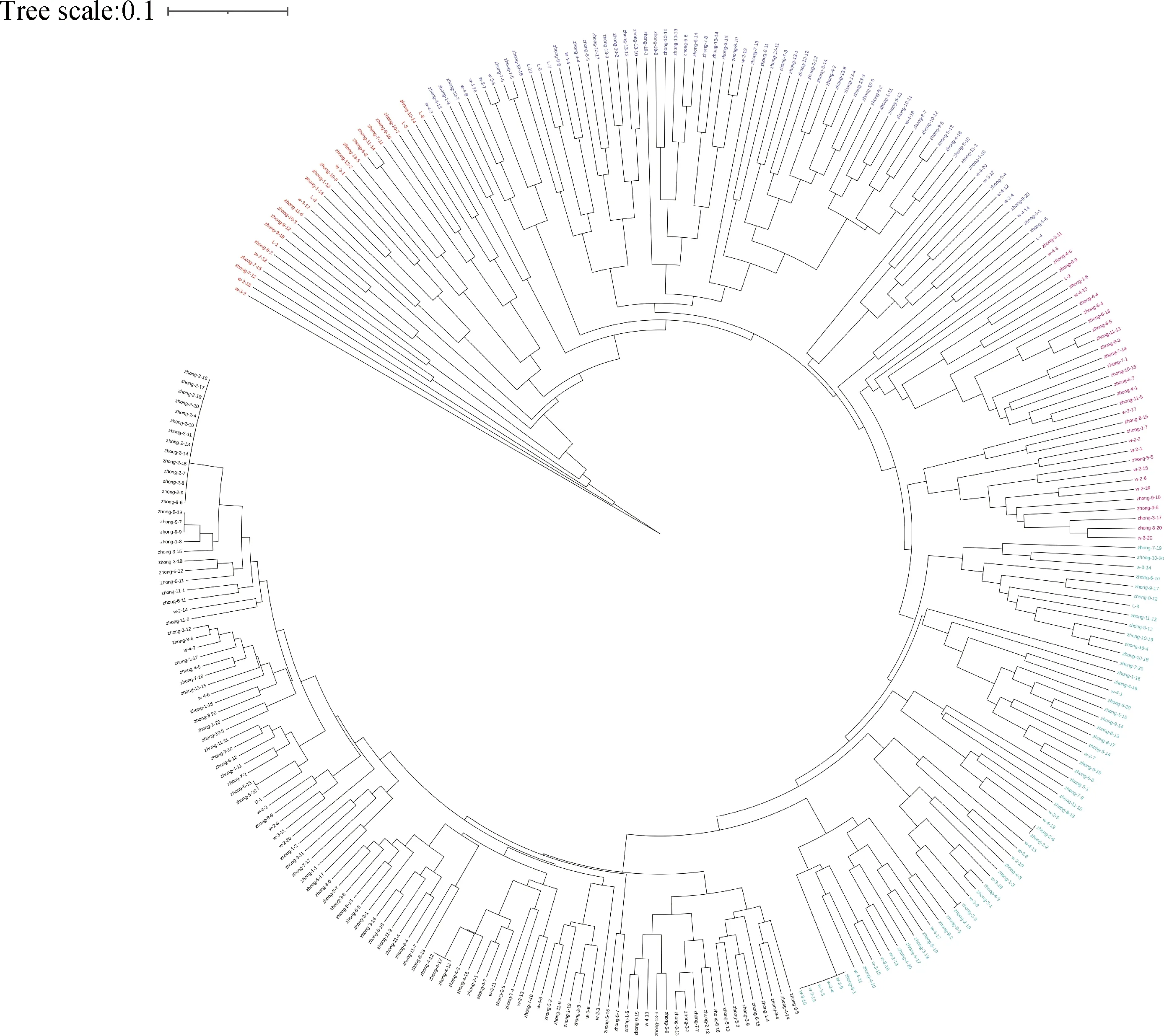
图5 299份红砂材料的聚类分析Fig.5 Clustering results of 299 R. Soongorica materials
3 讨论
3.1 居群遗传多样性
本研究的11对引物能成功扩增出3~9个等位基因,与张颖娟[10]和冯亮亮[14]的研究结果大致相同。等位基因多态率为83.58%,多态信息含量指数为0.395,得出红砂的基因位点具有较高的多态性,在今后的育种工作中可以利用该物种丰富的遗传材料,通过杂交、筛选等方法获得生态效益更高的红砂品种[27]。红砂居群的等位基因数均高于有效等位基因数,表明红砂的等位基因在染色体上分布不均匀,红砂居群在杂交过程中有明显的、通过染色体较大片段而进行遗传传递的现象。该现象可能会导致在遗传时发生后代遗传信息偏亲遗传的现象[28]。基因杂合度是衡量居群遗传变异的参数之一[29]。经研究表明红砂居群的杂合度均较低,其中居群MQXX的观测杂合度(0.286)最低,表明该居群内杂合率低,在杂交过程中产生的杂合度丢失的现象[30],推测产生该现象的原因与红砂的繁殖特性有关,位于干旱、沙化生境的成熟阶段的红砂居群主要以无性繁殖为主[3],而该红砂分布区常年高温少雨、干旱[31],因此居群内主要以根基劈裂和不定根的方式进行繁殖,使红砂居群内杂合度缺失及分化程度降低,进而导致对生存环境的适应性降低。本研究表明地方内红砂居群有一定程度的异交,这种交配所产生的后代具有杂合基因型。杂合基因型是产生基因交换、重组和产生新基因型的基础,这种交配方式的存在,提高了红砂居群内的基因杂合度,在红砂的进化过程中为其创造更广泛的遗传变异材料[32-33]。分化系数是从两个地方群体中任意抽取两个配子是同源的概率,可以度量红砂居群间的遗传分化程度[34]。结果表明红砂居群间的遗传分化主要来源于个体内DNA碱基序列的差异,AMOVA分析表明居群间遗传变异仅占遗传总材料的5.8%,这种结果与前人研究结果一致[10,35]。
3.2 遗传结构
本研究表明WD和SMY的遗传距离最近(0.007),MQXX和HG的遗传距离最远(0.229),Mantel检测得出红砂居群的遗传距离和地理距离显著相关(R2=0.3254,P=0.02<0.05),由此类现象推测出可能随着物种的进化所产生的地理隔离现象,经历不同的自然选择、不同的进化方向、进而产生种内差异而形成的地理小种[36-37]。聚类分析表明,来自腾格里沙漠东南边沿的SSC,SGK,WD,SMY,THCY,BYHT,NMYH,MJW,QYS被聚在第一类,这些居群在地理位置上较为集中,并且在红砂散粉期[38](7-8月)受中国夏季风的影响[39](风向由南向北,为夏季风盛行期),导致这些居群间有较频繁的基因交流,这种现象表明临近红砂居群间可以通过风媒促进居群间的基因交流,从而改变居群内遗传结构及富集基因库。来源于祁连山中段的LZ居群在遗传结构上其他居群分化明显,推测该现象是由于地理分布不同引起的种间差异[40],郝晓莉等[41]的研究结果表明五柱红砂(R.kaschgaricaRupr.)分布延伸至祁连山中段,因此LZ和其他居群的差异极有可能是由属内的不同种间引起的。居群QYS和MJW间的基因交流较频繁、遗传分化较小,极有可能是居群间能够自由交配的两个亚群,基因流在居群间是随机的、均一的,使得居群间相似性变大,进而导致居群间表现为基因频率和基因型频率的哈迪-温伯格平衡[42]。基因流弱化了自然选择和遗传漂变的作用,使居群间趋于一致,更有助于红砂群体内部能够稳定的生存和发育[43]。引物不能很好的区分THCY和BYHT,想要进一步了解红砂居群的遗传多样性及亲缘关系,需开发鉴别性更高的引物或在此基础上增加其他引物[44]。通过种质资源的遗传多样性分析,可有效降低资源利用过程中的盲目性。基于不同居群遗传结构表明,XGG,HSZ与其他居群遗传距离相对较远,居群内遗传多样性丰富,在进行红砂杂交组合选择时,可利用这2个居群的材料与其他材料进行组配,从而为育种工作者亲本选配提供科学依据[45]。
4 结论
本研究运用SSR分子标记技术,对16个不同地理来源的红砂居群进行遗传多样性及遗传结构分析。研究结论得出红砂的基因多态性为中度,等位基因不均匀的分布在染色体上。观测杂合度和F指数说明居群内杂合度缺失,纯合体过量。按照香农多样性指数评估红砂居群遗传多样性水平:THCY,ZZG>LZ>NMYH>QYS>HSZ>WWYH>MJW>SSC>BYHT>HG>XGG>SGK>WD>SMY>MQXX。Fst指数表明居群间分化程度较低,且红砂的遗传变异主要来自源于个体内DNA序列的不同。各居群间基因交流为3.163,说明居群间有较频繁的基因交流。Mantel检测得出遗传距离和地理距离显著相关。UPGMA将16个红砂居群分为5大类,聚类分析表明居群间的基因交流是影响居群聚类的重要因素。STRUCTURE结构分析将材料分为5个亚群。各亚群中,第Ⅴ亚群内的红砂个体血统较为纯正,第Ⅱ亚群内的红砂个体血统混杂。
