福建天宝岩不同长苞铁杉林土壤微生物生物量碳含量的比较及其与土壤含水量的关系
2022-09-27黄雄俊刘君成温鑫鸿游巍斌巫丽芸刘进山蔡昌棠何东进
黄雄俊,刘君成,温鑫鸿,游巍斌,巫丽芸,刘进山,蔡昌棠,何东进,3,①
(1.福建农林大学林学院,福建 福州 350002;2.福建天宝岩国家级自然保护区,福建 永安 366032;3.福建农业职业技术学院,福建 福州 350119)
森林土壤碳库对维持土壤生态系统的碳循环具有重要作用[1],其中土壤微生物生物量碳(microbial biomass carbon,MBC)作为土壤生态系统中最为活跃的有机质组分之一,其对林下微环境的变化十分敏感,既是森林植被在土壤中有效养分的重要储备库和来源[2,3],也是土壤有机质分解和养分转化过程的调控者[4]。因此,研究土壤微生物生物量碳的动态特征,不仅能了解土壤碳含量的季节变化,分析土壤健康状况,还能准确反映不同经营措施对土壤自然肥力和林分潜在生产力的提升程度[5,6]。
在森林植物群落中,优势树种由于一些自然或偶然原因死亡后,在树冠层中形成了空窗,即林窗[7],这是森林中普遍发生的自然现象,也是一种小规模的干扰形式。林窗会通过改变林内的光照、空气及土壤的温度、水分含量等环境条件,增大森林在空间分布上的不均匀性和复杂性,进而影响幼苗期植被的生长和更新速度以及土壤的养分物质循环[8-11]。李苏闽等[12]发现,在以长苞铁杉〔Nothotsugalongibracteata(W.C.Cheng)Hu ex C.N.Page〕为建群种的针叶林中,与非林窗区相比,林窗区的微环境因子变化幅度较小,且较之土壤温度和湿度变幅剧烈的大林窗,小林窗内微环境因子更为稳定,改善林窗内小气候的强度更高。费菲等[13]指出,在侧柏〔Platycladusorientalis(Linn.)Franco〕人工林中,相较于大林窗(450 m2)和小林窗(50 m2),中林窗(200 m2)对提高土壤微生物生物量碳和微生物生物量氮含量的作用更为明显,且能显著提升夏季林窗内土壤微生物生物量碳和微生物生物量氮含量,从而改善森林植被在生长季的生长状况。土壤中碳和氮的数量与土壤微生物生物量碳和微生物生物量氮的浓度密切相关,在不同林型中,因群落结构、生境条件、凋落物组分及分解难易程度等存在差异,土壤微生物生物量碳和微生物生物量氮含量及其动态特征也存在差异[14,15];不同林型形成的林窗在形状、面积、形成木及形成原因等变量间具有一定差异性,会引起林窗内光照、气温、土壤含水量及pH值等微环境条件的改变,进而影响土壤微生物群落的种类、数量及活性等,最终影响土壤微生物生物量碳的浓度及其动态变化[16,17],其中,土壤含水量是导致土壤微生物生物量碳含量差异的主要因子[18]。
长苞铁杉林是天宝岩国家级自然保护区森林的主体[19],拥有维持区域生态系统平衡、涵养水源和保持水土等重要生态作用[20]。长苞铁杉作为生长缓慢的喜阳树种,其幼苗在生长过程中难以与针阔叶树种竞争,且由于保护区内的长苞铁杉多为成熟或过熟林,立地条件相对贫瘠,长苞铁杉林正处于衰退状态,森林生态系统的服务功能难以正常发挥[21,22],如何以科学合理的途径来恢复其天然更新、增强其生态效应与生物多样性的保护功能成为亟待解决的问题。基于以上研究,综合考虑林型和林窗大小,本文对福建天宝岩国家级自然保护区4种典型长苞铁杉林的土壤微生物生物量碳含量及其与土壤含水量的相关性进行了分析,从土壤微生物生物量的角度探讨长苞铁杉不同林型和林窗大小对土壤养分循环过程的影响,以期为天宝岩国家级自然保护区长苞铁杉林的保护与合理经营提供科学依据。
1 研究区概况和研究方法
1.1 研究区概况
天宝岩国家级自然保护区位于福建省永安市东部,地跨青水、上坪和西洋3个乡(镇),具体地理坐标为东经117°28′03″~117°35′28″、北纬25°50′51″~26°01′20″,最高海拔1 604.8 m,最低海拔580 m,属中亚热带海洋性季风气候,年平均气温为15 ℃,7月平均气温为23 ℃,1月平均气温为5 ℃,无霜期约290 d,年平均降水量为2 039 mm,主要集中在夏、秋两季。保护区的占地面积达11 015.38 hm2,其中林地总面积为10 663.33 hm2,大面积分布的天然林构建了闽江地区重要的水源涵养林和生态屏障。保护区内植物种类丰富、结构复杂,主要的乔木树种有长苞铁杉、青冈(QuercusglaucaThunb.)、马尾松(PinusmassonianaLamb.)和毛竹〔Phyllostachysedulis(Carriere)J.Houzeau〕等;主要的灌木树种有猴头杜鹃(RhododendronsimiarumHance)和柃木(EuryajaponicaThunb.)等。森林类型分别以长苞铁杉和猴头杜鹃(冠幅大、树高较高,视为小乔木[23])为建群种的针叶林及山地苔藓矮曲林为主,二者构成的大面积原始森林群落皆是天宝岩国家级自然保护区内最具代表性的保护对象,尤其是珍稀濒危植物长苞铁杉林在保护区内的分布面积达186.7 hm2,是全国范围内面积最大的长苞铁杉纯林,极具保护和研究价值。
1.2 研究方法
在前期调查的基础上[22],于天宝岩国家级自然保护区海拔1 200~1 600 m范围内分散选取4种典型的长苞铁杉林(长苞铁杉纯林、长苞铁杉-青冈混交林、长苞铁杉-猴头杜鹃混交林和长苞铁杉-毛竹混交林),采取特定样方调查法,在同一长苞铁杉林林分中各取海拔高度相对一致、其他立地条件相似的大、中、小3种林窗(将长苞铁杉纯林中的3种林窗视为对照组),各林窗的基本特征见表1。

表1 福建天宝岩国家级自然保护区内不同长苞铁杉林12个林窗的基本特征
由于保护区内不同长苞铁杉林的树种组成复杂,优势树种高度与冠幅相差较大,且所处环境多为坡度较大的山地,导致在林冠层形成的林窗大小不一,因此,依据前期实地调查数据,将林窗按照面积(S)分为大(S>200 m2)、中(100 m2 参照段文标等[15]的研究方法,在12个林窗的中心区域使用罗盘仪确定正东、正西、正南、正北4个方向后用测绳连接至林窗边缘处(测绳区域为扩展林窗区),并从林窗边缘延长至非林窗区域。在中心区域设置1个面积2 m×2 m小样方,在扩展林窗区和延长线上的非林窗区内分别每隔1 m设置4个面积2 m×2 m和4个面积10 m×10 m的小样方,在每个林窗样地的9个样方中各随机设置1个取样点并标记1、2、3、4、5、6、7、8、9号,其中,1至5号取样点为林窗区,6至9号取样点为非林窗区。采样时小心移去取样点表层未分解的自然凋落物,用土钻取0~10 cm土层的土壤样品,其中1至5号取样点的样品混合为1个样品,6至9号取样点的样品混合为1个样品,分别装入自封袋并标记后带回实验室。共采样4次,分别为2019年5月、7月、9月、11月的中旬(即在春季、夏季、秋季和冬季分别采样)。 根据文献[15]中的方法测定土壤微生物生物量碳含量;采用烘干法[25]测定土壤含水量。 使用EXCEL 2021软件对实验数据进行整理和绘图;使用SPSS 21.0软件对实验数据进行单因素方差分析(one-way ANOVA),采用最小显著差异法(LSD)分析土壤微生物生物量碳在不同林型、林窗与非林窗及季节间的差异;采用Pearson相关性分析法分析土壤含水量与土壤微生物生物量碳间的相关性。 2.1.1 土壤微生物生物量碳含量的季节动态变化 结果(表2)显示:福建天宝岩国家级自然保护区4种长苞铁杉林林窗和非林窗内的土壤微生物生物量碳含量的季节差异总体达到了显著(P<0.05)水平,土壤微生物生物量碳含量的季节动态变化特征总体表现为春季和冬季高于夏季和秋季;相同季节同一林型大、中、小林窗与非林窗间的土壤微生物生物量碳含量差异总体达到了显著水平,与非林窗相比,大部分林窗内的土壤微生物生物量碳含量较高。 在长苞铁杉纯林中,各林窗内的土壤微生物生物量碳含量总体从高到低依次为冬季、春季、夏季、秋季,各林窗内的土壤微生物生物量碳含量在春季的差异最小。中、小林窗的非林窗内土壤微生物生物量碳含量的最小值均出现在秋季,分别为90.6和118.5 mg·kg-1。方差分析结果表明:长苞铁杉纯林林窗大小对土壤微生物生物量碳含量的影响不显著(P=0.35)。 在长苞铁杉-青冈混交林中,大、中、小林窗内的土壤微生物生物量碳含量的最大值均出现在春季,分别为397.2、396.4和273.8 mg·kg-1;最小值均出现在夏季,分别为134.3、167.5和102.2 mg·kg-1。各林窗内的土壤微生物生物量碳含量在秋季的差异最小。中、小林窗的非林窗内土壤微生物生物量碳含量的最小值均出现在秋季,分别为128.7和91.1 mg·kg-1;大、小林窗的非林窗内土壤微生物生物量碳含量的最大值均出现在春季,分别为243.3和191.7 mg·kg-1。方差分析结果表明:长苞铁杉-青冈混交林林窗大小对土壤微生物生物量碳含量的影响不显著(P=0.46)。 在长苞铁杉-猴头杜鹃混交林中,大、中、小林窗内的土壤微生物生物量碳含量的最大值均出现在春季,分别为341.5、272.0和461.8 mg·kg-1;大、小林窗内的土壤微生物生物量碳含量的最小值均出现在秋季,分别为121.5和159.7 mg·kg-1。各林窗内的土壤微生物生物量碳含量在秋季的差异最小。大、中林窗的非林窗内土壤微生物生物量碳含量的最大值均出现在冬季,分别为218.0和203.3 mg·kg-1。方差分析结果表明:长苞铁杉-猴头杜鹃混交林林窗大小对土壤微生物生物量碳含量的影响不显著(P=0.26)。 在长苞铁杉-毛竹混交林中,小林窗的非林窗内土壤微生物生物量碳含量从高到低依次为冬季、春季、夏季、秋季,其他大、中、小林窗及其非林窗内的土壤微生物生物量碳含量从高到低依次为春季、夏季、冬季、秋季。各林窗内的土壤微生物生物量碳含量在冬季的差异最小。方差分析表明:长苞铁杉-毛竹混交林林窗大小对土壤微生物生物量碳含量的影响不显著(P=0.59)。 2.1.2 土壤微生物生物量碳含量在不同林型间的比较 结果(表2)显示:在福建天宝岩国家级自然保护区,各林窗内土壤微生物生物量碳含量在不同林型间存在显著(P<0.05)差异。与长苞铁杉纯林相比,长苞铁杉-青冈混交林春季和秋季的大、中林窗内的土壤微生物生物量碳含量较高,长苞铁杉-猴头杜鹃混交林春季和夏季的大、小林窗内的土壤微生物生物量碳含量较高;长苞铁杉-青冈混交林秋季中林窗(229.8 mg·kg-1)内的土壤微生物生物量碳含量显著增加,长苞铁杉-猴头杜鹃混交林的春季小林窗(461.8 mg·kg-1)、夏季大林窗(319.4 mg·kg-1)和秋季中林窗(171.9 mg·kg-1)内的土壤微生物生物量碳含量显著增加,长苞铁杉-毛竹混交林冬季小林窗(174.7 mg·kg-1)内的土壤微生物生物量碳含量显著降低,其他林型各林窗内土壤微生物生物量碳含量与长苞铁杉纯林无显著差异。 表2 福建天宝岩国家级自然保护区不同长苞铁杉林土壤微生物生物量碳含量的比较 结果(表3)显示:总体上看,福建天宝岩国家级自然保护区相同季节同一林窗内的土壤含水量在不同林型间差异显著(P<0.05),长苞铁杉-青冈混交林和长苞铁杉-猴头杜鹃混交林林窗内的土壤含水量明显高于长苞铁杉纯林和长苞铁杉-毛竹混交林。 表3 福建天宝岩国家级自然保护区内不同长苞铁杉林林窗的土壤含水量比较 相同季节同一林型的土壤含水量在不同林窗间总体差异显著,春季和秋季中林窗内的土壤含水量明显高于大、小林窗,夏季大林窗内的土壤含水量明显高于中、小林窗,冬季长苞铁杉纯林和长苞铁杉-猴头杜鹃混交林大林窗以及长苞铁杉-青冈混交林和长苞铁杉-毛竹混交林中林窗内的土壤含水量较高。 对福建天宝岩国家级自然保护区不同长苞铁杉林土壤微生物生物量碳与土壤含水量进行相关性分析,结果显示:长苞铁杉纯林和长苞铁杉-青冈混交林林窗内的土壤微生物生物量碳含量与土壤含水量间均无显著相关性,相关系数分别为0.329和0.521;长苞铁杉-猴头杜鹃混交林和长苞铁杉-毛竹混交林林窗内的土壤微生物生物量碳含量与土壤含水量分别呈极显著(P<0.01)和显著(P<0.05)负相关,相关系数分别为-0.788和-0.638。表明在长苞铁杉-猴头杜鹃混交林和长苞铁杉-毛竹混交林的林窗内,土壤微生物生物量碳含量与土壤含水量密切相关,土壤含水量的变化能够影响土壤微生物生物量碳含量。 土壤微生物生物量碳的季节动态变化较为复杂且存在不确定性,受林内植被类型、干扰类型和程度、空气和土壤的温度、水分含量以及pH值等多种环境因子的影响[26-28]。因此,在不同林型中,土壤微生物生物量碳的季节动态变化有所不同,且在同一林型中,即使环境因子相同,不同植被类型下土壤微生物生物量碳的季节动态变化也可能存在差异[29]。周义贵等[30]发现,在米亚罗林区云杉(PiceaasperataMast.)低效林中,表层(0~15 cm)和亚表层(15~30 cm)的土壤微生物生物量碳含量均在植物生长季末期达到峰值,而在植物生长旺季最小,由高至低依次为秋季、冬季、春季、夏季。本研究中,4种长苞铁杉林林窗与非林窗内的土壤微生物生物量碳含量的季节差异总体达到了显著水平,土壤微生物生物量碳含量的季节动态变化特征总体表现为春季和冬季高于夏季和秋季(“夏低冬高”[5])。在福建天宝岩国家级自然保护区内,春季适宜的水热条件为土壤微生物提供良好的生长环境,增强了微生物的生物活性并加快其物质和能量代谢,使土壤微生物生物量碳含量增加;夏季和早秋土壤微生物生物量碳含量较少可能是由于高温、高湿的环境条件加快了植物的生长发育,导致植物对土壤中碳、氮等养分的需求增大,且立地条件和资源受限也影响了土壤微生物的大量繁殖,进而影响土壤的微生物生物量[5,30],还可能是由于进入雨季后,降水量增大的同时也增强了雨水的淋溶作用,有效养分的流失使土壤微生物生物量减少;冬季植物进入生长季末期或休眠期,生长速度减缓,根系吸收的养分含量降低[3],土壤微生物生物量碳在土壤中得以积存。 林窗的形成改变了林内光照的空间分布格局,对土壤中微生物的周转速率及其代谢活性产生影响,而微生物又通过分解过程在碳素循环中作用于土壤生态系统[31]。在费菲等[13]的研究中,侧柏人工林中林窗(200 m2)内的土壤微生物生物量碳、微生物生物量氮明显高于大林窗(450 m2)和小林窗(50 m2);肖建强等[32]指出,在辽东山区次生林处于发育后期的林窗中,大(670 m2)、中(290 m2)、小(90 m2)3种林窗间的土壤微生物生物量碳、微生物生物量氮和微生物生物量磷均无显著差异。本研究结果表明:与非林窗相比,大部分林窗内的土壤微生物生物量碳含量较高,且总体达到显著水平,而大、中、小林窗间的土壤微生物生物量碳含量总体差异不显著。说明林窗的形成对提升土壤微生物生物量碳含量有一定的显著影响,但林窗大小对土壤微生物生物量碳含量的影响并不显著。相关研究结果表明:由于在林窗对土壤微生物生物量碳的影响过程中受土壤理化性质[33]、植物种类[29]、底物有效性[30]、枯倒木剩余生物量[34]等因子的综合影响,在林窗形成初期,空气和土壤中的温度、水分含量发生的有效性变化会显著影响土壤微生物生物量碳含量,但随着林窗年龄的增加以及林内天然更新形成幼林,林窗区与非林窗区的生境条件逐渐趋于一致,林窗内的土壤养分状况也逐渐恢复至郁闭林分水平[35],这可能是导致本研究不同林窗间无显著差异的主要原因。相关性分析结果表明:在长苞铁杉-猴头杜鹃混交林和长苞铁杉-毛竹混交林的林窗内,土壤微生物生物量碳含量与土壤含水量分别呈极显著和显著负相关,表明林窗中较高的土壤含水量会在一定程度上限制土壤微生物生物量碳含量的增加,该结果与前人研究结论[16,30]基本一致;而在长苞铁杉纯林和长苞铁杉-青冈混交林的林窗内土壤微生物生物量碳含量和土壤含水量间均无显著相关性,这可能是由于不同林型土壤微生物生物量碳含量增加的主要限制因子不同[13,35]。 本研究中,相较于长苞铁杉纯林,长苞铁杉-青冈混交林春季和秋季的大、中林窗内的土壤微生物生物量碳含量较高,长苞铁杉-猴头杜鹃混交林春季和夏季的大、小林窗内的土壤微生物生物量碳含量较高,且这2种长苞铁杉混交林林窗内的土壤含水量也较高,表明在福建天宝岩国家级自然保护区植物生长旺季,以长苞铁杉为优势树种分别形成的长苞铁杉-青冈混交林和长苞铁杉-猴头杜鹃混交林对林窗内土壤微生物生物量碳含量的提高效果较为明显,这可能是由于较之纯林,混交林林内光照梯度变化明显、物种构成复杂以及适宜水热条件形成良好的林内小气候,促进了土壤微生物的能耗和新陈代谢,提高了土壤微生物生物量碳含量[36-38];而长苞铁杉-毛竹混交林各林窗内的土壤微生物生物量碳含量总体较低,这主要是由于与竹针混交林相比,长苞铁杉-青冈混交林和长苞铁杉-猴头杜鹃混交林林下凋落物总量及其组分类型丰富,腐殖质层厚且水分含量高[39,40],有利于森林植被的更新生长,为土壤微生物提供了适宜的成长环境,加快凋落物的分解速度,从而增加了土壤微生物生物量碳含量。 本文仅探究了福建天宝岩国家级自然保护区不同长苞铁林林窗内表层(0~15 cm)土壤的微生物生物量碳含量及其与土壤含水量的关系,对于土壤微生物活性、更深层的土壤微生物生物量碳含量及其他土壤微生物生物量(氮、磷等)等问题缺乏必要的研究,难以全面评价林窗对土壤微生物生物量碳含量的影响。在今后的研究中还应考虑将土壤微生物群落、土壤表层和亚表层(15~30 cm)等对微生物生物量的影响进行综合比较和分析,进一步完善林窗对地上和地下养分循环影响的研究。 综上所述,福建天宝岩国家级自然保护区4种长苞铁杉林林窗和非林窗内的土壤微生物生物量碳含量的季节差异总体达到了显著水平,林窗与非林窗间的土壤微生物生物量碳含量差异也总体达到了显著水平,与非林窗区相比,大部分林窗内的土壤微生物生物量碳含量较高,而大、中、小林窗间的土壤微生物生物量碳含量总体差异不显著。相较于长苞铁杉纯林和长苞铁杉-毛竹混交林,长苞铁杉-青冈混交林和长苞铁杉-猴头杜鹃混交林林窗内的土壤微生物生物量碳含量和土壤含水量较高。因此,长苞铁杉-青冈混交林和长苞铁杉-猴头杜鹃混交林以及林窗的形成均能较好地改善林内小气候,为长苞铁杉群落和土壤微生物提供更适宜的生境条件,从而提高长苞铁杉林生态系统的生产力和稳定性,并促进保护区内长苞铁杉林的土壤碳循环过程。1.3 数据处理
2 结果和分析
2.1 不同长苞铁杉林土壤微生物生物量碳含量的比较
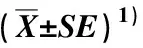
2.2 不同长苞铁杉林林窗内土壤含水量的比较
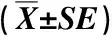
2.3 不同长苞铁杉林土壤微生物生物量碳含量与土壤含水量的相关性
3 讨论和结论
