二纺蛛总科(蜘蛛目:新蛛下目)蜘蛛化石研究进展
2022-09-26郭向博任东
郭向博,任东
首都师范大学生命科学学院,北京,100048
内容提要:二纺蛛总科包括5个现生科、5个化石科以及一些科级位置未定的化石物种,是蜘蛛目中较为古老的一个类群。目前已知最早的二纺蛛总科化石记录可以追溯到早侏罗世,但分子系统发育研究推测其起源时间为二叠纪。二纺蛛总科蜘蛛在中生代最为繁盛,化石物种相对较多,是蜘蛛化石类群的重要组成部分。而现生二纺蛛总科蜘蛛十分稀少,且主要分布在热带和亚热带地区。近年来的系统发育研究认为二纺蛛总科可能是复杂生殖器类的姐妹群,是蜘蛛演化研究的关键类群。因此,本文对二纺蛛总科分类系统的研究历史进行了回顾,总结了二纺蛛总科化石类群在不同地质历史时期的组成和分布,统计了二纺蛛总科蜘蛛化石的名录。目前,二纺蛛总科蜘蛛化石分类研究基础薄弱,部分化石类群的系统位置存疑,二纺蛛总科内部系统发育关系尚不清晰。在将来的研究中,需要利用多种技术手段获取化石材料的形态学信息,打好分类学基础;结合化石类群和现生类群进行系统发育分析;进而联系古地理、古生态、生物学行为等研究,全方位分析二纺蛛总科的自然演化历史。
二纺蛛总科Palpimanoidea隶属于蜘蛛目Araneae后纺亚目Opisthothelae新蛛下目Araneomorphae,是蜘蛛目中的重要类群。现生二纺蛛总科蜘蛛十分稀少,目前已报道291种,主要分布在热带和亚热带地区(World Spider Catalog, 2022),均为捕食性,捕食行为复杂多样。并且,大多数现生二纺蛛总科蜘蛛具有“食蛛性”(araneophagy),专性或偏好捕食其他蜘蛛,并演化出了特殊的适应性结构,如头区向上强烈延伸、螯肢延长、背甲和第一对步足表皮增厚、第一对步足膨大等(Pekár et al., 2011; Wood et al., 2012; Platnick, 2020)。
二纺蛛总科最古老的化石记录可以追溯到早侏罗世(Selden and Dunlop, 2014),包括5个现生科(古蛛科Archaeidae、胡通蛛科Huttoniidae、展颈蛛科Mecysmaucheniidae、二纺蛛科Palpimanidae和斯坦蛛科Stenochilidae)和5个化石科(蜥蛛科Lagonomegopidae、小二纺蛛科Micropalpimanidae、平古蛛科Planarchaeidae、斯贝蛛科Spatiatoridae和维蒂蛛科Vetiatoridae)。在中生代,二纺蛛总科物种数约占同时代蜘蛛目物种总数的27%,是蜘蛛化石类群的重要组成部分(Dunlop et al., 2020)。并且,近年来的系统发育研究认为二纺蛛总科可能是复杂生殖器类Entelegynae的姐妹群(Wood et al., 2012, 2018; Wheeler et al., 2017; Fernández et al., 2018),复杂生殖器类是现生蜘蛛中多样性最高的类群(约占总物种数量的80%)。所以,二纺蛛总科的起源与演化在整个蜘蛛目的演化历程中有十分重要的意义。本文对二纺蛛总科蜘蛛化石研究现状进行概述,并对存在的问题和研究前景加以分析。
1 二纺蛛总科分类系统的研究简史
二纺蛛总科蜘蛛的分类学研究起始于1820年,Dufour(1820)报道了首个二纺蛛总科蜘蛛Palpimanusgibbulus,建立二纺蛛属Palpimanus,但当时处于分类学发展的初期,总科这一阶元的概念尚未出现。随着进化思想和系统学观念的兴起,分类学中用到的高级阶元也越来越多。Levi(1982)将横螯类(Labidognatha)蜘蛛分为16个总科,其中二纺蛛总科包含4个科:拟平腹蛛科Zodariidae、展颈蛛科、二纺蛛科和斯坦蛛科,鉴别特征为后纺器退化或缺失。然而Levi(1982)的分类体系并未得到其它学者的认可(Platnick, 1983)。Forster 和 Platnick(1984)利用比较形态学和支序学的方法对古蛛科及其相关类群进行了研究,提出了包含10个现生科的二纺蛛总科分类系统,即二纺蛛总科包含古蛛科、展颈蛛科、二纺蛛科、斯坦蛛科、胡通蛛科、拟古蛛科Pararchaeidae、全古蛛科Holarchaeidae、塔克蛛科Textricellidae、小幽蛛科Micropholcommatidae和拟态蛛科Mimetidae,它们的共有衍征为螯肢具腺体丘,以及前齿堤具钉状齿。随后,Platnick和Forster(1986, 1987)对自己于1984年提出的分类系统进行了补充和修订,取消塔克蛛科的科级地位,认为塔克蛛科是小幽蛛科的异名,并且将马尔卡蛛科Malkaridae归入二纺蛛总科。
21世纪以来,随着支序学和分子系统发育的应用越来越广泛,Forster和Platnick(1984)的分类系统受到了越来越多的质疑。通过对拟古蛛科等7个科的形态特征进行对比,Schütt(2000)认为应该将塔克蛛科和小幽蛛科从二纺蛛总科转移至园蛛总科Araneoidea,拟态蛛科和马尔卡蛛科也很可能属于园蛛总科。Griswold 等(2005)基于形态学和行为学数据,对复杂生殖器类蜘蛛进行了系统发育分析,结果表明Forster和Platnick(1984)的二纺蛛总科不是单系群,拟态蛛科和拟古蛛科很可能属于园蛛总科。另外,一些基于分子数据的系统发育分析也支持将小幽蛛科、拟古蛛科、全古蛛科和拟态蛛科从二纺蛛总科中移除(Rix et al., 2008; Dimitrov et al., 2012)。Wood 等(2012)结合形态数据和分子数据,对古蛛科(包括现生类群和化石类群)及其相关类群进行了系统发育分析,认为二纺蛛总科应包含5个现生科:古蛛科、展颈蛛科、二纺蛛科、斯坦蛛科和胡通蛛科,这个分类系统被其他蛛形学者广泛接受并沿用至今。
在化石研究方面,Petrunkevitch(1942)基于波罗的海琥珀(Baltic amber)标本建立的斯贝蛛科,是第一个被归入二纺蛛总科的化石科。Forster和Platnick(1984)在讨论中引用了Wunderlich的观点:由于步足具抹刀状刚毛,斯贝蛛科可能应该被归入二纺蛛总科。随后,Wunderlich(1986)将斯贝蛛科正式归入二纺蛛总科。Eskov 和Wunderlich(1995)报道了4个保存在俄罗斯泰梅尔琥珀(Taimyr amber)中的幼蛛标本,并据此建立了蜥蛛科,蜥蛛科区别于蜘蛛目中其他科的特征是:背甲前端两侧具一对巨大的后中眼。Eskov和Wunderlich(1995)根据螯肢前齿堤具钉状齿、步足跗节无听毛、以及步足无刺等特征,将蜥蛛科归入了二纺蛛总科。
白垩纪缅甸琥珀中保存了大量精美的节肢动物化石,Wunderlich是对缅甸琥珀蜘蛛化石研究贡献最大的蛛形学者之一。他报道了大量保存在缅甸琥珀中的蜘蛛化石,并且据此建立了小二纺蛛科、维蒂蛛科和平古蛛科,三者都被归入了二纺蛛总科(Wunderlich, 2008a, 2012, 2015, 2017a; Wunderlich and Müller, 2021)。目前,二纺蛛总科包含5个现生科(古蛛科、展颈蛛科、二纺蛛科、斯坦蛛科和胡通蛛科)和5个化石科(蜥蛛科、斯贝蛛科、小二纺蛛科、维蒂蛛科和平古蛛科)。除此之外,二纺蛛总科还包含一些科级位置未定的化石类群,共有4属5种(Selden and Dunlop, 2014; Selden et al., 2008, 2020)。
2 二纺蛛总科蜘蛛化石的地质历史
二纺蛛总科的化石类群包括8科43属86种,以及科级位置未定类群4属5种(图1;表1)。目前已知最古老的二纺蛛总科化石记录可以追溯到早侏罗世,但基于分子钟的研究推测二纺蛛总科蜘蛛的起源时间为二叠纪(Fernández et al., 2018; Magalhaes et al., 2020)。
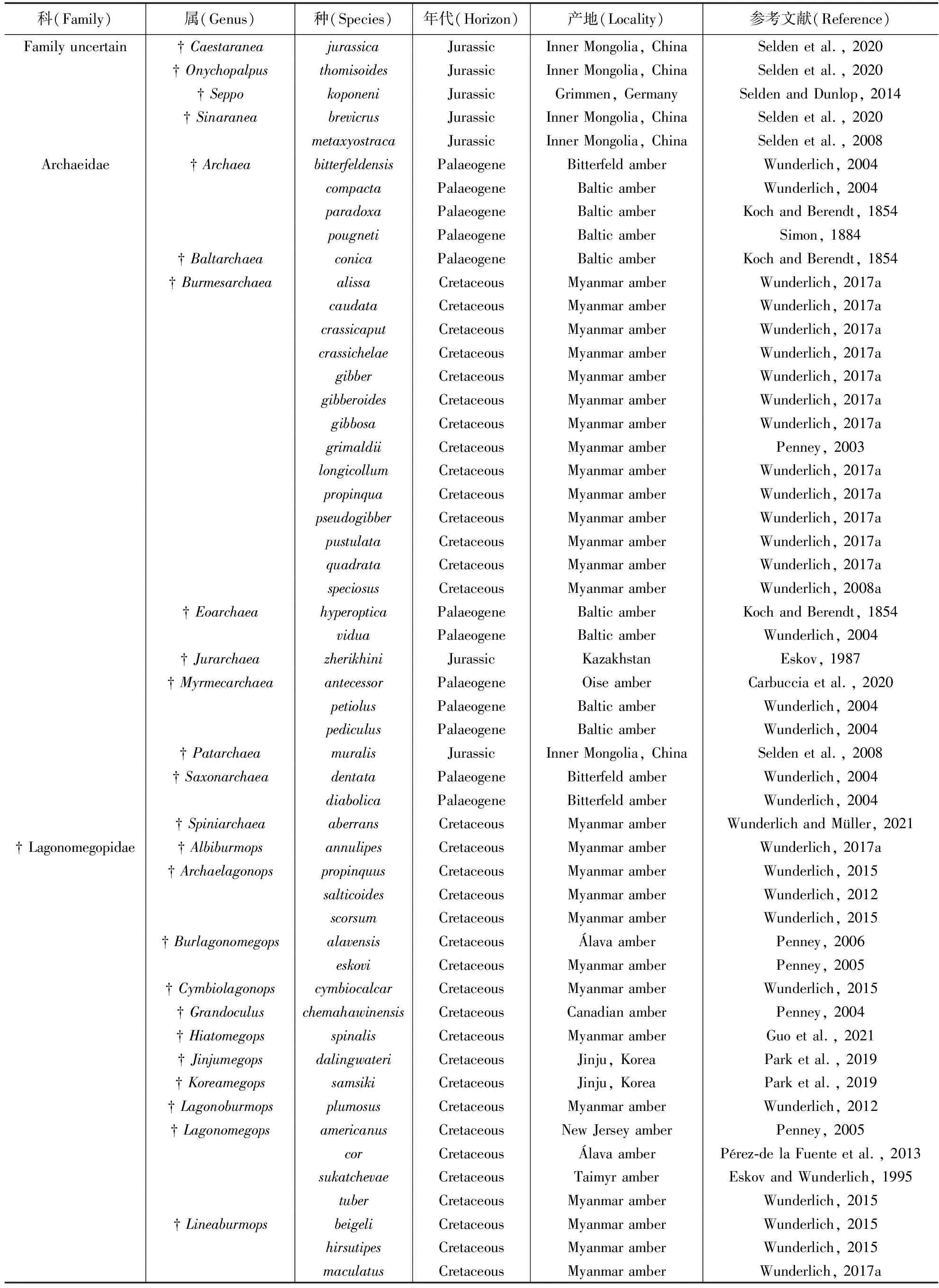
表1 二纺蛛总科蜘蛛化石名录

图 1 二纺蛛总科蜘蛛化石代表
2.1 侏罗纪二纺蛛总科蜘蛛
侏罗纪的二纺蛛总科蜘蛛化石数量较少,均为印痕化石和印模化石,包括古蛛科2属2种以及科级位置未定类群4属5种。产自德国格里门地区(Grimmen)下侏罗统地层的印模化石蜘蛛Seppokoponeni是目前已知最古老的二纺蛛总科化石物种,但由于保存较差,Seppokoponeni无法被归入特定的科(Selden and Dunlop, 2014)。位于中国东北部宁城县的道虎沟化石产地出土了大量精美的中侏罗世动植物化石,一直是古生物学研究的热点地区。Selden等(2008, 2020)报道了5个产于此地的二纺蛛总科化石物种,其中包括4个科级位置未定的化石物种和1个古蛛科化石物种Patarchaeamuralis,Patarchaeamuralis也是目前已知最古老的古蛛科化石记录。另外一个侏罗纪时期的古蛛科化石物种Jurarchaeazherikhini,产自哈萨克斯坦的上侏罗统地层(Eskov, 1987)。
2.2 白垩纪二纺蛛总科蜘蛛
白垩纪是二纺蛛总科化石蜘蛛最繁盛的时期,这一时期的二纺蛛总科蜘蛛化石的种类和数量都十分丰富,共有8科36属68种,并且一些化石科(蜥蛛科、小二纺蛛科、平古蛛科和维蒂蛛科)仅发现于白垩系地层中。白垩纪时期的二纺蛛总科蜘蛛化石主要保存在琥珀中,琥珀产地有西班牙(早白垩世)、约旦(早白垩世)、法国(早白垩世)、缅甸(晚白垩世早期)、俄罗斯泰梅尔(晚白垩世)、加拿大(晚白垩世)、美国新泽西(晚白垩世)。其中缅甸琥珀(Myanmar amber)是二纺蛛总科蜘蛛多样性最高的化石产地,目前已经有约60%的二纺蛛总科化石物种发现于此。除了琥珀化石,Park 等(2019)报道了两个产自韩国下白垩统地层中的蜥蛛科印痕化石物种Jinjumegopsdalingwateri和Koreamegopssamsiki;Downen 和 Selden(2021)描述了一个产自巴西下白垩统地层中的二纺蛛科印模化石物种Cretapalpusvittari,这也是目前二纺蛛科仅有的一个化石记录。另外,Penney和Selden(2006)报道了一些保存在晚白垩世加拿大琥珀(Canadian amber)中的蜘蛛化石,并根据背甲、眼列、步足刺、跗节爪、抹刀状毛等特征将其归入胡通蛛科,但由于标本均为幼蛛且保存较差,未进行正式命名。
2.3 古近纪二纺蛛总科蜘蛛
到了古近纪,园蛛总科和RTA分支蜘蛛类群开始兴盛(Magalhaes et al., 2020)。这一时期的二纺蛛总科蜘蛛化石记录相对较少,共有2科6属16种,全部保存在欧洲的琥珀中:波罗的海琥珀、德国比特费尔德琥珀(Bitterfeld amber)、法国瓦兹琥珀(Oise amber)。斯贝蛛科是二纺蛛总科中唯一一个从白垩纪延续至古近纪的化石科,在缅甸琥珀、波罗的海琥珀和比特费尔德琥珀中均有斯贝蛛科的化石记录(Petrunkevitch, 1942; Wunderlich, 2006, 2008b, 2015, 2017b)。
2.4 新近纪和第四纪二纺蛛总科蜘蛛
在新近纪和第四纪,二纺蛛总科蜘蛛仅在马达加斯加柯巴脂(Madagascar copal)中有过报道(Lourenço, 2000)。Lourenço(2000)描述了保存在马达加斯加柯巴脂中的一个古蛛科亚化石,将其命名为Archaeacopalensis,并指出新标本和现生物种Madagascarchaeagracilicollis十分相似。随后,该亚化石被认为是Madagascarchaeagracilicollis的幼蛛标本(Wunderlich, 2004)。柯巴脂是琥珀形成过程中的一个中间阶段,地质年代距今更近,里面保存的内含物也更加接近现在的生物类群。尽管在过去的20多年中,已经有一些研究报道了保存在马达加斯加柯巴脂中的节肢动物类群,但其地质年代一直存在争议(Poinar, 1999; Lourenço, 2000; Vitali, 2007; Bosselaers et al., 2010; Labandeira et al., 2014; Bukejs and Legalov, 2017)。最近的一项研究认为马达加斯加柯巴脂仅有几百年的历史,里面的内含物不具备传统的古生物学研究意义,但是在研究人类活动对生物多样性的影响时,马达加斯加柯巴脂可以发挥自己的作用(Delclòs et al., 2020)。
3 二纺蛛总科化石研究中存在的问题
(1)分类研究基础薄弱,化石鉴定困难。由于化石保存等原因,许多已报道的二纺蛛总科蜘蛛化石特征描述与绘图简单,所提供的形态学信息有限。这给后续的分类鉴定和系统发育工作增加了难度与挑战。另外,在二纺蛛总科中,一些化石类群是仅基于未成熟个体建立的,比如蜥蛛科至少有12个物种仅有幼蛛标本的记录和描述(Park et al., 2019)。幼蛛尚未发育完全,与成熟蜘蛛之间存在形态结构上的差异(Foelix, 2011)。因此,在进行分类研究时,幼蛛与成蛛不易正确匹配,容易造成同物异名或错误鉴定。
(2)部分类群系统位置存疑。一直以来,对蜘蛛系统发育关系的研究大多仅基于现生类群的形态、行为和分子数据,得到的结果也只能描绘现生类群间的系统发育关系(如Griswold et al., 2005; Bond et al., 2014; Fernández et al., 2014; Ramírez, 2014; Polotow et al., 2015; Garrison et al., 2016; Dimitrov et al., 2017; Wheeler et al., 2017; Azevedo et al., 2018; Wood et al., 2018; Opatova et al., 2020)。而化石类群通常被用来在系统发育树上标定特定节点的时间(如Dimitrov et al., 2013; Benavides et al., 2017; Fernández et al., 2018; Li Fengyuan and Li Shuqiang, 2018; Shao Lili and Li Shuqiang, 2018; Magalhaes et al., 2020),很少作为终端类群用于系统发育分析。因此,在蜘蛛目中,许多化石类群的系统位置存在争议(Magalhaes et al., 2020)。在二纺蛛总科的5个化石科中,只有蜥蛛科的系统位置得到了支序分析的验证。蜥蛛科在建立之初就被归入了二纺蛛总科,当时依据的特征是螯肢前齿堤具钉状齿,步足后跗节无听毛以及步足无刺(Eskov and Wunderlich, 1995)。但是,Eskov和Wunderlich(1995)描述并命名的首个蜥蛛物种的模式标本是幼蛛标本,尚未发育完全。随着越来越多蜥蛛化石(尤其是成熟个体)被报道,研究者们对蜥蛛的认识也越来越全面。Penney(2004, 2005)首次在蜥蛛科物种中报道了步足跗节听毛的存在,而在二纺蛛总科其他类群中,均未发现上述结构。Pérez-de la Fuente 等(2013)对比了蜥蛛科和二纺蛛总科中5个现生科的背甲、眼的排列方式、螯肢、颚叶、下唇、步足听毛以及步足特化形式等特征,认为将蜥蛛科归入二纺蛛总科是有待验证的。Guo Xiangbo等(2020; 2021)描述了一些未在蜥蛛科中报道过的新特征,并在此基础上将蜥蛛化石作为终端类群进行支序分析,结果支持把蜥蛛科归入二纺蛛总科,验证了蜥蛛科的系统位置。
(3)二纺蛛总科的单系性及内部系统发育关系尚不清晰。在Wood 等(2012)的研究中,基于全证据(形态数据+分子数据)和仅依靠形态数据的分析结果均支持他们新定义的二纺蛛总科为单系群,而仅依靠分子数据的分析结果不支持这个类群的单系性。随后的一些系统发育分析也在这个问题上存在争议(Wheeler et al., 2017; Fernández et al., 2018; Wood et al., 2018; Guo Xiangbo et al., 2021)。Wood 等(2018)利用目标序列捕获测序技术对馆藏蜘蛛标本的超保守元件(Ultra-Conserved Elements)和外显子位点(exonic regions)进行测序,并进行系统发育分析,重建了二纺蛛总科内部各现生科间的系统发育关系。这是近年来仅有的聚焦于二纺蛛总科内部系统发育关系的研究。但是,在上述的系统发育分析中,仅Wood等(2012)和Guo Xiangbo等(2021)的研究涉及部分古蛛科和蜥蛛科化石类群,其余分析皆是基于现生类群的分子数据进行的。二纺蛛总科内部化石类群与现生类群之间的系统发育关系仍未得到澄清。
4 二纺蛛总科化石研究展望
在漫长的地质历史中,二纺蛛总科这一相对古老的蜘蛛类群经历了由辉煌到衰落的过程。中生代是二纺蛛总科最繁盛的时期,物种数约占同时代蜘蛛目物种总数的27%,但其真实的多样性远不止于此。并且,在已发表的二纺蛛总科化石记录中,仍有许多物种的描述不够详尽,分类地位值得商榷。因此,利用多种技术手段(扫描电镜、Micro-CT成像及3D重建等)充分发掘二纺蛛总科蜘蛛化石的形态学信息,描述新物种并修订已报道物种仍是今后二纺蛛总科化石研究的重点。
二纺蛛总科可能是复杂生殖器类的姐妹群,探究二纺蛛总科系统发育关系意义重大。但近年来的系统发育分析结果未能在二纺蛛总科单系性的问题上达成一致。并且,由于缺乏对化石类群广泛取样的系统发育研究,二纺蛛总科内部化石类群与现生类群之间系统发育关系尚不清晰。通过古今结合、宏观与微观相结合的方式,包含全面的二纺蛛总科内部类群,基于充足的形态学数据和分子数据的系统发育分析亟待进行。此外,二纺蛛总科现生类群分布较窄,主要在热带和亚热带地区,而化石产地广泛分布于北半球。结合系统发育分析、古地理及大陆板块漂移学说进行历史生物地理学研究,推测二纺蛛总科蜘蛛的起源和分布格局的形成也会成为将来研究的重点内容。
部分现生二纺蛛总科蜘蛛具有特殊的食性(食蛛性)和捕食行为,这些行为与其适应性结构紧密联系。在地质历史背景下,对相关形态结构演化的研究可以反映出这些特殊行为的起源和演化历程,进而探讨二纺蛛总科蜘蛛演化与生态环境变化之间的关系,全方位分析二纺蛛总科的自然演化历史。这也是未来研究的一个重要方向。
谨以此文祝贺杨文采主编80华诞!
