不同密度下山桐子自然整枝差异性分析
2022-09-23朱秀征房丽莎王艳梅蔡齐飞耿晓东
朱秀征, 靳 超, 房丽莎, 刘 震, 王艳梅,蔡齐飞, 李 志, 耿晓东
(河南农业大学林学院,郑州 450002)
自然整枝(natural pruning)是指当林分郁闭达到一定程度后,各单木的生长空间受到限制,树冠下部的枝条因光照和营养供应不足而开始枯死和脱落的现象[1]. 自然整枝受遗传因素和环境的影响较大,目前其强度主要以树干直径、冠高比、枝下高、保留活枝数及枝条特征等指标来评判[2]. 密度对林木成活、成长、成林、成材和结果等都起着巨大的作用[3],随着林分密度的减小,林木的胸径和单株材积均会显著增加[4-5],还有研究表明林分密度对林木的冠幅、活枝下高、冠长和冠长率都有显著影响[6]. 林分密度不同,其自然整枝时间和强度均不同[7],高密度的林分枝条死亡速度大于低密度林分,有利于自然整枝减少死节的形成[8],而低密度的林分枝条生长空间更大,与相邻枝条竞争更小,自然整枝强度也更小[9]. 一般认为,越容易发生自然整枝的林木其枯枝直径也越细[2],目前,不少学者已经通过研究建立了枝条生长[10]、枝下高[11]以及树冠生长[12]等模型来揭示林木自然整枝的规律.
山桐子(Idesia polycarpa)是大风子科(Flacourtiaceae)山桐子属落叶乔木. 果实(浆果)含油率高且营养丰富,其种子、果肉与果实的含油率分别可达28.17%,48.35%与39.40%,油脂中的成分有亚油酸等十余种,且不同种源之间有差异[13]. 山桐子不仅是优质、高产的木本油料植物,同时还兼有较高的用材、观赏价值,被誉为“美丽的树上油库”[14]. 山桐子虽然在我国分布广泛,但多为野生、散生,呈零星分布,少有片林[15],且山桐子种源和家系之间各生长指标差异较大[16-17]. 目前,关于山桐子的研究多集中在实生苗培育[18]、油脂理化性质[19-20]等方面,密度及遗传因素是如何影响山桐子自然整枝的还未有学者进行系统的研究. 因此,本试验以三种密度下两个种源的5 a生山桐子幼树为试验材料,通过分析不同密度下山桐子自然整枝强度的差异以及各生长指标对自然整枝的影响,了解密度与自然整枝的关系,为通过密度调控和修枝措施来培育高质量的山桐子林提供理论依据和技术指导.
1 材料与方法
1.1 试验地概况
试验地位于河南农业大学科教园区(113°59′E、34°87′N),该地区属暖温带大陆性气候,年平均气温14.4 ℃,7月最热,月平均气温27.3 ℃,1月最冷,月平均气温0.2 ℃;年平均降雨量640.9 mm,无霜期220 d,全年日照时间约2400 h. 试验地土壤质地为砂壤土,土壤pH 为7.5,有机质含量为15.78 g/kg,碱解氮含量为54.24 mg/kg,速效磷含量为17.59 mg/kg,速效钾含量为143.51 mg/kg.
1.2 材料来源
试验材料为引种于河南济源(104°06′E,30°67′N)和四川成都(112°57′E,35°08′N)并栽培于河南农业大学科教园区长势良好的5 a 生山桐子幼树. 2011 年12 月分别在河南济源和四川成都采种,每个种源的种子均采摘于同一棵山桐子母树,2012 年3 月将经过低温处理的种子播种在河南农业大学科教园区,6 月选取地径和苗高一致的幼苗移栽至大田,定期对苗木进行除草和浇水的日常管理,不进行其他干预,使其自然生长,试验观测时两个种源山桐子的郁闭度均达到了0.8 以上,均未发生自然稀疏且长势良好,保存率在95%以上.
1.3 试验设计
采用双因素完全随机区组设计,成都种源和济源种源的山桐子各设置3种株行距处理,分别为T1:3 m×2 m、T2:3 m×3 m和T3:3 m×4 m,种源和株行距配置如图1 所示. 共6 种处理,每个处理3 次重复,共计18种处理,每个处理20株山桐子幼树,共计360株山桐子幼树.

图1 山桐子种源和株行距配置图Fig.1 The schematic diagram of provenance and plant row spacing configuration of I.polycarpa
1.4 观测内容和方法
2016 年6 月分别对每个处理的山桐子进行胸径、树高、枝下高以及冠幅的测量和单株材积的计算,然后分层统计第1轮、第2轮和第3轮枝条的长度和基径、枝条着生处主干高度和直径以及枝条死亡率. 胸径、枝条基径和枝条着生处主干直径用游标卡尺测量,精确到0.1 cm,树高、冠幅、枝条长度和着生处主干高度用卷尺测量,精确到0.1 m. 单株材积的计算采用平均实验形数法[19],如公式(1)所示,每个轮次枝条死亡率的计算方法如公式(2)所示. 图2为5 a生山桐子不同轮次示意图.


图2 山桐子不同轮次示意图Fig.2 The schematic diagram of different rounds in I.polycarpa
式中:V为单株材积;g1.3为胸径断面积;h为树高;f0为实验形数,阔叶树一般为0.40.

1.5 数据处理与分析
利用WPS Office 软件进行数据整理,Origin 2017 绘图,并使用SPSS24.0软件对不同处理下山桐子的胸径、枝下高、冠高比、单株材积、枝条长度与基径、枝条着生处主干高度与直径以及枝条死亡率进行单因素方差分析. 若差异显著,则用邓肯多重比较法在0.05水平下进行多重比较,之后采用P=0.05和P=0.01的皮尔逊相关性分析得出山桐子各个生长指标与自然整枝的相关性.
2 结果与分析
2.1 密度对不同种源山桐子胸径、枝下高、冠高比及单株材积的影响
由表1可以看出,成都种源山桐子在不同密度下胸径、冠高比和单株材积均表现为T2 >T3 >T1,但各处理间差异均不显著(P>0.05);济源种源山桐子在不同密度下胸径和单株材积均表现为T3>T2>T1,其中T2和T3处理下的胸径和单株材积显著大于T1处理(P<0.05),胸径和单株材积最大分别为3.7 cm和0.002 6 m3,分别是T1 处理的2.6 和6.5 倍. 两个种源山桐子的枝下高在不同密度下差异均不显著(P>0.05),而冠高比在不同密度下均表现为T2 最高. 成都种源山桐子的枝下高和单株材积在3个密度下均显著高于济源种源(P<0.05),成都种源山桐子的胸径在T1和T2处理下显著高于济源种源(P<0.05),但在T3处理下的胸径与济源种源差异性不显著(P>0.05),两个种源山桐子的冠高比在3个密度下差异性均不显著(P>0.05).
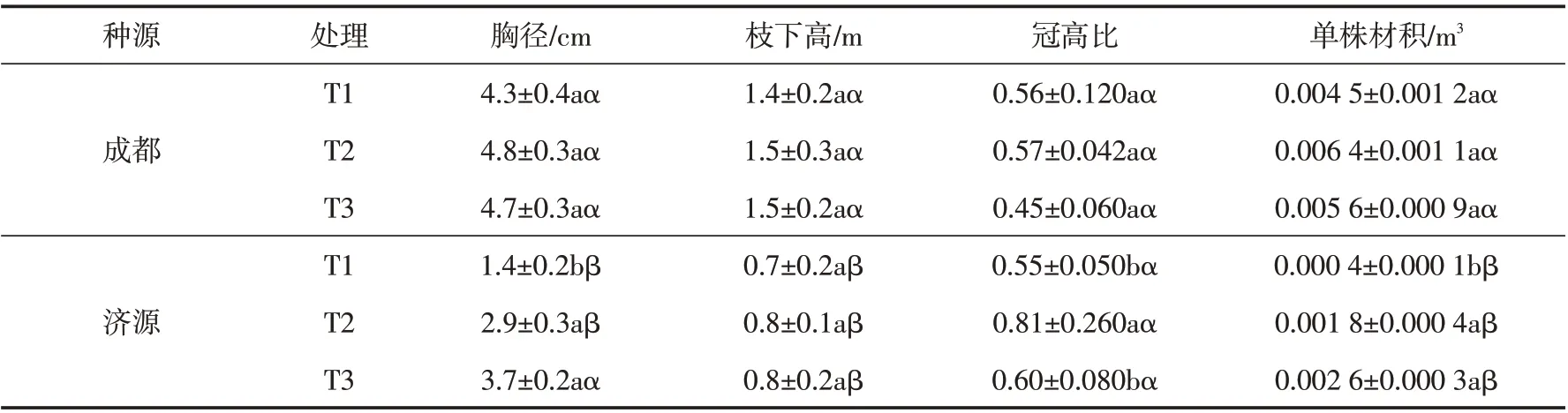
表1 不同密度下成都和济源种源山桐子的胸径、枝下高、冠高比和单株材积Tab.1 The DBH,under-branch height,crown height ratio and individual volume of I.polycarpa from Chengdu and Jiyuan under different densities
2.2 密度对不同种源山桐子枝条生长的影响
由图3可知,成都种源山桐子3个轮次的枝条长度在不同密度下表现为T3>T2>T1,而济源种源山桐子的枝条长度则为T2处理大于其他两个处理,两个种源山桐子的枝条长度和枝条基径在不同密度下均没有表现出显著的差异性(P>0.05);两个种源山桐子3个轮次枝条的死亡率在不同密度下均表现为T1>T2>T3,在相同密度下均表现为第1轮>第2轮>第3轮. 总体来看,成都种源山桐子的枝条长度和枝条死亡率在3种密度处理下均显著大于济源种源(P<0.05).
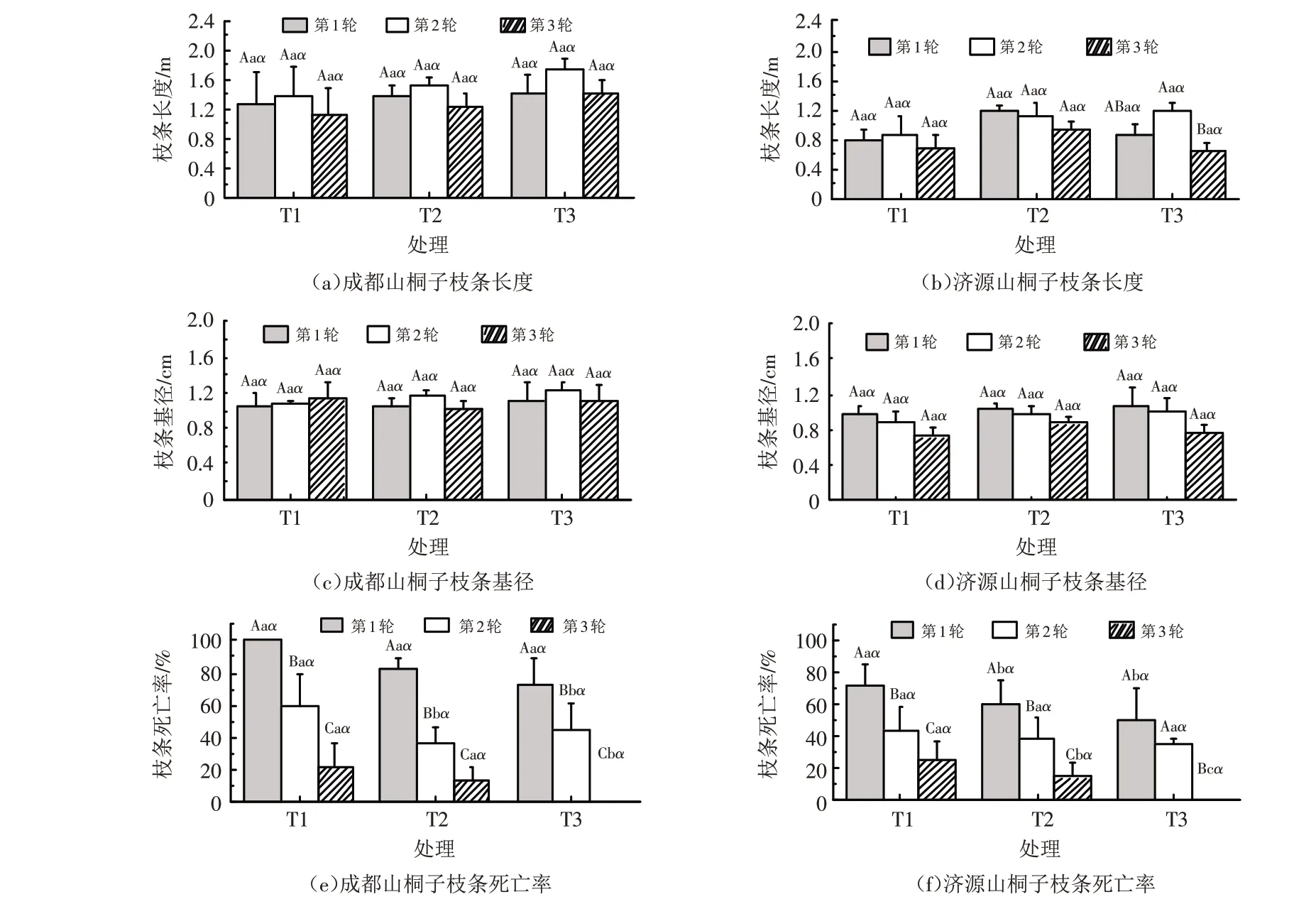
图3 不同密度下两个种源山桐子枝条长度、枝条基径和枝条死亡率Fig.3 The branch length,branch diameter and branch mortality of I.polycarpa from two provenances under different densities
2.3 不同栽植密度对不同种源山桐子枝条着生位置的影响
由图4 可知,两个种源山桐子的3 个轮次的枝条着生处主干直径在不同密度下均表现为T3>T2>T1,其中,济源种源山桐子T1处理下第1轮和第2轮枝条的着生处主干直径显著小于T2和T3处理(P<0.05);两个种源山桐子的3个轮次的枝条着生处主干高度在3个密度下差异性均不显著(P>0.05). 整体来看,成都种源山桐子3个轮次的枝条着生处主干直径和着生处主干高度也大于济源种源.
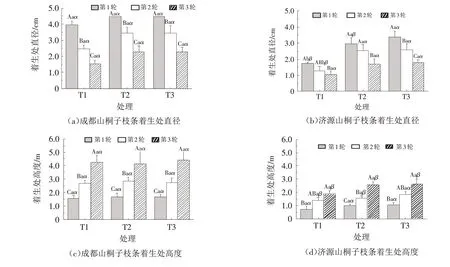
图4 不同密度下两个种源山桐子枝条着生处主干直径和着生处主干高度Fig.4 The diameter and height of branches’site in I.polycarpa from two provenances under different densities
2.4 两个种源山桐子各生长指标与自然整枝的相关性分析
由表2 可知,成都种源山桐子的第1 轮枝条死亡率与第1 轮枝条的长度和基径有极显著的负相关关系(P<0.01). 济源种源山桐子第1轮枝条死亡率,与第1轮枝条着生处主干直径、第2轮枝条着生处主干直径、第2轮枝条长度、第3轮枝条长度以及第3轮枝条着生处主干直径均有显著或极显著的相关性;第2轮枝条死亡率,与第1轮枝条长度、第1轮枝条着生处主干直径、第2轮枝条着生处主干直径、第3轮枝条着生处主干直径以及第3 轮枝条长度有显著或极显著的相关性;第3 轮枝条死亡率,与第1 轮枝条着生处主干直径、第2轮枝条着生处主干直径、第3轮枝条着生处主干直径有显著或极显著的相关性.

表2 成都种源和济源种源山桐子各生长指标与自然整枝的相关性分析Tab.2 The correlation analysis between growth index and natural pruning of I.polycarpa from Chengdu and Jiyuan
3 讨论
通常情况下,在林木生长过程中,林木的树干直径、枝下高、冠高比和单株材积等可以在一定程度上影响其自然整枝发生的时间和强度. 童书振等[21]发现杉木(Cunninghamia lanceolata)人工林分经密度调控后,低密度经营有利于林分的胸径、树高、单株材积加快生长,但其自然整枝能力受到了削弱[22]. 在本研究中,随着密度的减小,两个种源山桐子的胸径和单株材积均有所增加,而其自然整枝强度则显著减小,与前人研究结果一致. 一般来说,林木的枝下高与其自然整枝能力有极显著的相关关系,枝下高越高代表其自然整枝强度越高[23],但在本研究中不同密度下两个种源山桐子的枝下高均没有显著差异,自然整枝强度与枝下高的相关性也不显著,可能是用于试验的山桐子林分还未完全郁闭,自然整枝强度较小. 随着整枝强度的减小,林木的冠幅会有所增大,冠高比则会显著增加[24],但是在本研究中,随着两个种源山桐子的自然整枝强度减小,山桐子的冠高比却表现为先增大后减小,与前人的研究结果不一致,可能是由于当林分密度过小时,5 a生山桐子的树冠开张已经达到了极致,而树高则受密度的影响逐渐增大,导致了山桐子冠高比的减小.
枝条的大小和数量决定着生叶片的数量,叶片数量越多,便可以产生越多的光合产物用于枝条本身的发育[25],从而减少枝条的死亡. 王俊杰[26]发现不同造林密度下2 a 生中华猕猴桃(Actinidia chinensis)的枝条长度表现为低密度大于高密度,可能是因为高密度的林分光环境较弱,枝条的伸长生长受到了抑制[27].在本研究中,两个种源山桐子的枝条长度均表现为密度越大,枝条长度越小,与前人的研究结果一致;与枝条长度不同,两个种源山桐子的枝条基径在不同密度下基本不变,而王春胜[28]通过研究发现树冠内所有枝条平均直径均随着密度的增加呈现显著降低的趋势. 其他学者对桉树(Eucalyptus robusta)[29]以及花旗松(Pseudotsuga menziesii)[30]的研究结果也是如此,可能是由于山桐子在幼树时期通过光合作用产生的有机物主要用于枝条的伸长生长,导致山桐子幼树枝条基径的变化不大. 除此之外,密度越小,自然整枝强度越小,两个种源山桐子的枝条着生处主干直径越大. 这种现象可以在前人对桉树的研究中证实:随着密度的减小,桉树的冠幅增加,可以积累更多的光合产物,因此树干的断面积会持续增加,树冠基部也会逐渐后退从而导致枝条着生处主干直径的增加[31]. Hein等[32]在建立不同株行距下挪威云杉(Picea asperata)分支特性模型时也发现树枝存活的概率与其在树枝轮内的相对直径有关.
遗传因素对不同种林木的自然整枝均有不同程度的影响,许忠海等[33]发现不同种源山杨(Populus davidiana)的胸径、侧枝粗、树干通直度和自然整枝能力等均有显著或极显著的差异性,不同种源山桐子的胸径、树高及枝上高等生长指标的年生长规律均存在显著差异[34]. 从本研究结果可以看出,两个种源山桐子的各生长指标及自然整枝强度在不同密度下均表现为成都种源高于济源种源,可能是因为成都种源山桐子在前期生长较快,林分郁闭程度更大才导致了其自然整枝强度更高.
4 结语
5 a生山桐子在3个密度下均发生自然整枝现象,密度可以通过影响山桐子的胸径、冠高比及枝条发育状况和枝条着生位置来影响其自然整枝. 受遗传因素的影响,在同一密度下成都种源山桐子自然整枝强度显著大于济源种源. 因此在山桐子林的营造过程中,应通过合理的密度调控和人工修枝措施来减小其自然整枝强度,从而达到相应的造林目的.
