鸟类的兼性低体温
2022-09-06轩辕玉洁柳劲松
轩辕玉洁,柳劲松
鸟类的兼性低体温
轩辕玉洁,柳劲松
(温州大学 生命与环境科学学院,浙江 温州 325035)
鸟类是恒温动物,可以通过自身产生大量的热以维持体温的恒定.然而,自然界中有些鸟类在面临能量需求增加时,可以通过降低体温的方式减少能量消耗以维持生存.鸟类的兼性低体温是指鸟类静止时的身体核温显著低于正常活动状态下鸟类的体温,包括静止低体温、日蛰伏和休眠.影响鸟类兼性低体温的因素包括环境温度、食物、激素水平和繁殖周期等.对鸟类兼性低体温生存策略进行简述,旨在为后续研究提供参考.
鸟类;兼性低体温;环境温度;食物
鸟类是恒温动物,通过内源性代谢产热来维持较高的体温.鸟类的高体温暗示了其巨大的能量支出,即代谢产热相对较高.然而,鸟类的一些行为和生理机制可以降低其代谢水平,包括生境的选择、群居、兼性低体温以及代谢特征的适应性变化,如银喉长尾山雀()可以利用兼性夜间低温来降低低环境温度下的能量消耗[1].目前,采用无线电遥感勘测法研究自由放养鸟类的体温调节模式发现,白颊黑雁()、波多短尾鴗()、澳洲裸鼻鸱()、白背鼠鸟()等多数种类在夜间出现低体温现象[2].这些行为和生理上的适应性变化通常使得鸟类的代谢及体温具有明显的节律,活动时体温较高,静止时体温较低[3]706.近年来的研究表明,鸟类兼性低温发生在更多的鸟类中,对鸟类的能量平衡发挥着重要作用[4-6].
1 鸟类的兼性低体温
Reinertsen[7],McKechnie[3]等将鸟类夜间体温明显低于白天正常活动水平定义为兼性低体温,分为深度低体温(休眠及日蛰伏)和浅度低体温(静止低体温).日蛰伏与休眠之间的区别在于其持续的长度及最低的临界体温,日蛰伏与静止低体温之间的界线尚不清晰.目前,公认的标准是体温下降幅度大于10 ℃即认为鸟类休眠或日蛰伏,相反则认为是静止低体温[3]706.以往研究表明,仅体重低于80 g的小型鸟类出现日蛰伏和休眠现象,而Körtner[8]等发现,体重约500 g的茶色蟆口鸱()在冬季也能休眠,表明出现兼性低体温的鸟类的体重已经超过了先前的预测范围.Reinertsen[7]认为鸟类在夜间出现的兼性低体温反应可能是作为应对食物短缺或冷环境压力的一种策略,对其降低体温调节能力或减少能量消耗具有重要的作用.然而,对黑帽山雀()的研究表明,兼性体温过低也可能发生在白天,这种体温过低的发生可能是个别山雀选择的结果,且并不妨碍它们积极觅食[9].
鸟类与哺乳类同属于恒温动物,但这两类动物对外部刺激所做出的兼性低体温反应有所不同.Geiser[10]等根据恒温动物低体温持续时间的长度、最低临界体温以及代谢减少程度等生理参数对动物表现出来的兼性低体温反应进行分类,并认为大多数情况下,鸟类低体温反应的生理参数与哺乳类的日蛰伏和休眠类似.然而这些鸟类与哺乳类不同,其体温虽低于正常水平,下降幅却不超过10℃,只表现出静止低体温或受调控的低体温.大多数具有休眠和日蛰伏特征的哺乳动物其体温调节遵循“进入-维持-苏醒”模式.在进入阶段,其主要特征是代谢下调和随后的体温下降;维持阶段则是对某一特定体温值的调节;苏醒阶段动物的代谢产热增加,使得体温上升到正常水平.与哺乳类相比,鸟类在兼性低体温期间体温调节模式是相对可变的,在不同种类之间和不同个体之间可能存在大量不同的体温调节模式[11].如虹彩蜂鸟()、黑蜂鸟()、燕尾蜂鸟()在半自然条件下体温调节模式存在多种形式(见图1).
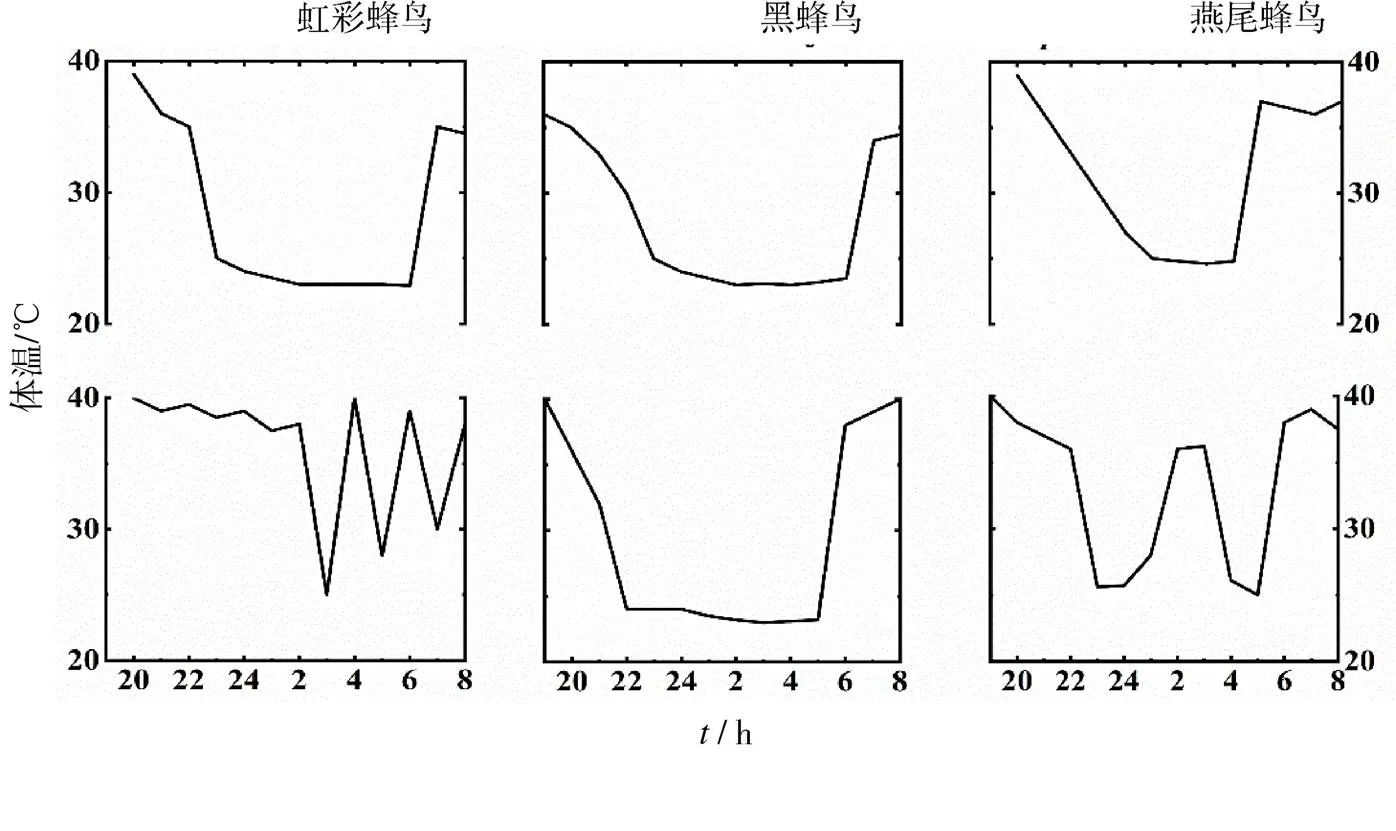
图1 3种蜂鸟在半自然条件下的体温调节模式[3]
2 鸟类兼性低体温的地理分布
在所有的动物地理区系中,除了东洋界以外均有报道一些鸟类物种的兼性低体温反应(见表1),随着纬度从高到低,鸟类出现低体温的物种数有增加的趋势,如澳大利亚界11种,古北界16种,新热带界29种.
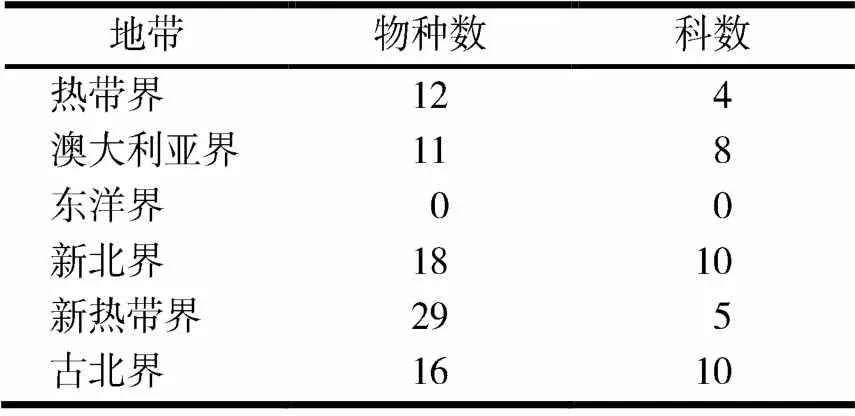
表1 鸟类兼性低体温的地理分布[3]
在鸟类的11个目中,部分种类曾报道存在兼性低体温,但大多数鸟类未发现有低体温现象[3]714-715.对部分鸟类的体温数据(ΔT/℃=norm-min.式中:ΔT为体温变化幅;norm为正常体温;min为最低临界体温)进行了分类统计(见表2).由表2可见,鸟类的兼性低体温能力表现出较大的种属差异性,且受体重影响较大,如体型较大的雁鸭类和雉鸡类的体温变化幅度均小于10 ℃[12-13].此外,更明显的低体温(ΔT≥20 ℃)仅在雨燕科、蜂鸟科和夜鹰科等少数科中发现.

表2 鸟类兼性体温分类统计(ΔTb/℃=Tnorm-Tmin)
3 鸟类兼性低温起源及影响因素
关于鸟类兼性低体温的起源,Geiser[13]认为鸟类兼性低体温与较小的体型和不可预测的食物供给联系密切,但Malan[14]认为鸟类的兼性低体温可能与物种的进化有关,如哺乳动物中相对原始的物种象鼩()具有明显的日蛰伏直接证明了该假说,红背鼠鸟()和班鼠鸟()的体温调节类型也为该假说提供了间接支持[3]716.在系统发育较为原始的类群中,鸟类的兼性低体温与当前关于动物异温性的演变观点相符合.
环境温度是导致鸟类兼性低体温的因素之一.大多数小型鸟类在低温条件下表现出不同程度的兼性低体温反应.不同程度的低体温在生理上表现为代谢率的变化,鸟类低体温时的代谢率与正常活动时的代谢率有显著差异[15].如相比于日间,鸠鸽科(Columbidae)小长尾鸠()夜间体温为34.4 ℃,降低4 ℃,代谢率减少22.0%;燕科(Hirundinidae)毛脚燕()夜间体温为25.7 ℃,降低13.2 ℃,代谢率减少20.0%;夜鹰科(Eurostopodidae)的夜鹰()夜间体温为29.6 ℃,降低10.9 ℃,代谢率减少51.8%等[16].这些鸟类代谢率下降程度直接反映了低体温期间节约能量的大小.
食物是引起鸟类夜间体温降低的另一重要因素.作为应对夜间食物摄入量减少的一种策略,改变体温调节能力或减少能量消耗具有重要作用.Reinertsen[7]认为鸟类在夜间出现的兼性低体温反应可能是作为应对食物短缺或冷环境压力的一种策略,对其增加体温调节能力或减少能量消耗具有重要作用[3,17].褐头山雀()在限食条件下体温会降低5.9 ℃,崖沙燕()在食物减少时体温降低5.1 ℃,而长时间饥饿导致仓鸮()自身能量储备不足,最终在夜间出现低体温反应[18].早期的研究表明,食物是引起宽尾煌蜂鸟()日蛰伏唯一、直接的决定因子[19].如宽尾煌蜂鸟在食物充足的时候采取不蛰伏的对策,而在夜间食物匮乏时则有蛰伏现象(见图2).后期更深入的研究表明,蜂鸟蛰伏受一系列因子决定,包括季节性变化、感知食物的可利用性及同化率的暂时性变化等[20].鸟类在食物短缺时立即进入低体温状态使其能量支出达到最小,以减少食物短缺带来的生存压力.
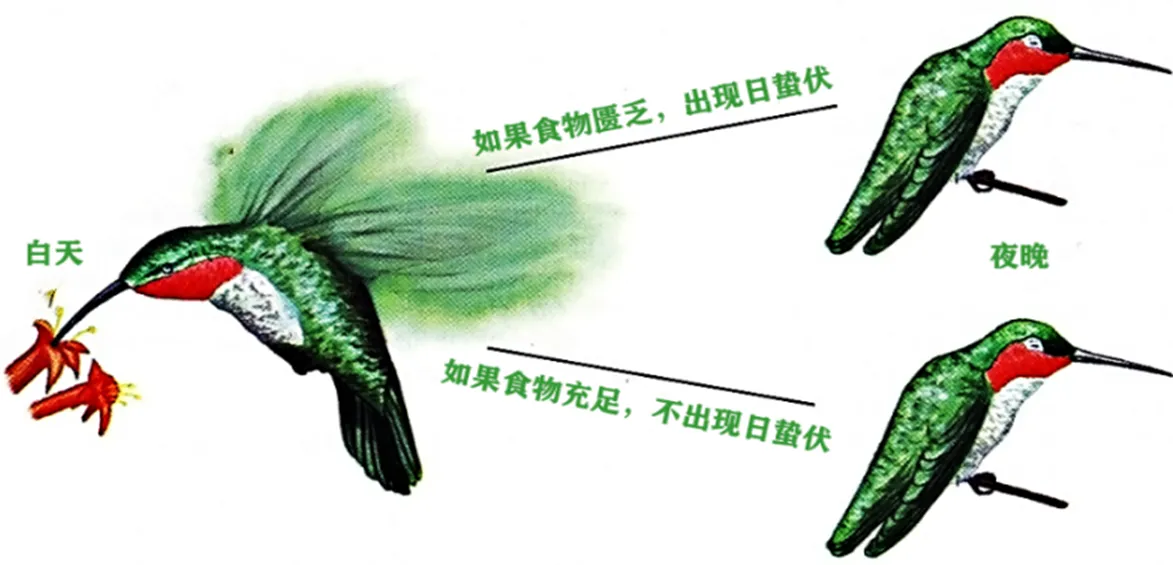
图2 食物决定宽尾煌蜂鸟是否进入日蛰伏[21]
为了降低被捕食的风险并且度过夜间食物短缺的时段,小型鸟类常常需要调节夜间体温以节省能量储备[4]919.如当蓝山雀()的脂肪储备充足时会维持正常体温,但体内脂肪储备不足会引起体温降低[22].鸟类能量储备的增加,一方面能提高生存和繁殖成功率,另一方面也是应对夜间不可预测的食物获取的一种生存策略,其最终目的是为了躲避危险以及增加代谢产热,保持巢内温度,确保卵孵化成功[23].
繁殖周期和激素水平在鸟类的兼性低体温中起到重要作用.Merola-Zwartjes[2]等发现,波多黎各短尾鳼()的休眠仅限于雌性动物,表明性激素可能对不同性别鸟类的休眠产生影响.雄性走鹃()在繁殖期可以维持正常体温,而雌性和非繁殖期间的雄性依然存在低体温的现象[24];繁殖期的蓝鹱()即使在低温环境或食物短缺时也不出现低体温现象.与此类似,繁殖期的小型哺乳类动物的雄性激素水平可能是其日蛰伏和冬眠的影响因素,如去雄的南非囊鼠()易进入蛰伏,通过注射雄性激素后则可抑制蛰伏[25].
鸟类兼性低体温是一种长期进化、发展、演化而来的生存策略,受环境温度、食物资源、激素水平及繁殖周期等一系列的生理生态因素共同调节,能够有效地应对食物短缺或低温环境压力,在减少能量消耗方面具有重要作用.我国地处东洋界,至今未见该地域鸟类出现深度低体温现象报道,未来研究前景广阔.
[1] Hatchwell B J,Sharp S P,Simeoni M,et al.Factors influencing overnight loss of body mass in the communal roosts of a social bird[J].Functional Ecology,2009,23(2):367-372.
[2] Merola-Zwartjes M,Ligon J D.Ecological energetics of the Puerto Rican Tody:heterothermy,torpor and intra-island variation[J].Ecology,2000,81(4):990-1003.
[3] McKechnie A E,Lovegrove B G.Avian Facultative Hypothermic Responses:A Review[J].Condor,2002,104(4):705-724.
[4] Brodin A,Nilsson J K,Nord A.Adaptive temperature regulation in the little bird in winter:predictions from a stochastic dynamic programming model[J].Oecologia,2017,185(1):1-12.
[5] Broggi J,Koivula K,Hohtola E,et al.Environmental drivers of roosting body mass variation in boreal Great Tits[J].Ibis,2017,159(4):919-924.
[6] Geiser F.Ecological physiology of daily torpor and hibernation[M].Gewerbestrasse:Springer,2021.
[7] Reinertsen R E.Physiological and ecologicalaspects of hypothermia,In CareyC(ed),Avian energetics and nutritional ecology[M].New York:Chapman and Hall,1996,125-157.
[8] Körtner G,Brigham R M,Geiser F.Winter torpor in a large bird[J].Nature,2000,407(6802):318.
[9] Lewden A,Petit M,Milbergue M,et al.Evidence of facultative daytime hypothermia in a small passerine wintering at northern latitudes[J].Ibis,2014,156(2):321-329.
[10] Geiser F,Ruf T.Hibernation versus dailytorpor in mammals and birds:physiological variables and classification of torpor patterns [J].Physiological Zoology,1995,68(6):935-966.
[11] Shankar A,Cisneros INH,Thompson S,et al.A heterothermic spectrum in hummingbirds[J].Journal of Experimental Biology,2022;225(2):jeb243208.
[12] Prothero J,JÜrgens K D.An energeticmodel of daily torpor in endotherms[J].Journal of Theoretical Biology,1986,121(4):403-415.
[13] Geiser F.Evolution of daily torpor and hibernation in birds and mammals:importance of body size[J].Clinical & Experimental Pharmacology & Physiology,1998,25(9):736-740.
[14] Malan A.The origins of hibernation:a reappraisal,In Geiser F,Hulbert A J,and Nicol S C(eds),Adaptations to the cold:Tenth International Hibernation Symposium[M].Armidale:University of New England Press,1996,1-6.
[15] Maddocks T A,Geiser F.Energetics,thermoregulation and nocturnal hypothermia in Australian Silvereyes[J].Condor,1997,99(1):104-112.
[16] Prinzinger R,Siedle K.Ontogeny of metabolism,thermoregulation and torpor in the House Martin(L.)and its ecological significance[J].Oecologia,1988,76(2):307-312.
[17] Kuepper N D,Carina M,Nestor C,et al.Facultative hypothermia as a survival strategy during snowstorm induced food shortages in Antarctic storm-petrel chicks[J].Comparative Biochemistry and Physiology:Part A Molecular and Integrative Physiology,2018,224:76-83.
[18] Thouzeau C,Duchamp C,Handrich Y.Energy metabolism and body temperature of barn owls fasting in the cold[J].Physiological and Biochemical Zoology,1999,72(2):170-178.
[19] Hainsworth F R,Collins B G,Wolf L L.The function of torpor in hummingbirds[J].Physiological Zoology,1977,50(3):215-222.
[20] McWhorter T J,Martĺnezdel Rio C.Does gut function limit hummingbird food intake[J].Physiological and Biochemical Zoology,2000,73(3):313-324.
[21] Molles Jr M C,Sher A A.Ecology:Concepts and Applications(Eighth edition)[M].New York:McGraw-Hill Education,2019,122.
[22] Nord A,Nilsson J F,Nilsson J A.Nocturnal body temperature in wintering Blue Tits is affected by roost-site temperature and body reserves[J].Oecologia,2011,167(1):21-25.
[23] Macleod R,Barnett P,Clark J A,et al.Body mass change strategies in blackbirds:The starvation-predation risk trade-off[J].Journal of Animal Ecology,2005,74(2):292-302.
[24] Vehrencamp S L.Body temperatures of incubating versus non-incubating roadrunners[J].Condor,1982,84(2):203-207.
[25] Mzilikazi N,Lovegrove B G.Reproductive activity influences thermoregulation and torpor in pouched mice,[J].Journal of Comparative Physiology B,2002,172(1):7-16.
Facultative hypothermia in birds
XUANYUAN Yujie,LIU Jinsong
(School of Life and Environmental Sciences,Wenzhou University,Wenzhou 325025,China)
Birds are homeotherm which can produce a lot of heat by itself to maintain a constant body temperature.When faced with increased energy demands, some birds in nature can reduce energy consumption by lowering their body temperature to maintain survival.Facultative hypothermia is defined when birds body temperature is significantly below the active level,including rest-phase hypothermia,daily torpor and hibernation.There are many factors influencing bird hypothermia response,including environmental temperature, food,hormone level and reproductive cycle.Reviews the survival strategy of facultative hypothermia in birds,aiming to provide reference for subsequent research.
bird;facultative hypothermia;ambient temperature;food
1007-9831(2022)08-0082-05
Q95
A
10.3969/j.issn.1007-9831.2022.08.016
2022-05-06
国家自然科学基金资助项目(31971420)
轩辕玉洁(1996-),女,安徽合肥人,在读硕士研究生,从事动物生理生态学研究.E-mail:20451334035@stu.wzu.edu.cn
柳劲松(1964-),男,黑龙江齐齐哈尔人,教授,博士,从事动物生理生态学研究.E-mail:ljs@wzu.edu.cn
