基因编辑技术在蔬菜中的研究进展
2022-08-27唐笑王桂香刘凡韩硕宗梅郭宁段蒙蒙
唐笑 王桂香刘凡 韩硕 宗梅 郭宁 段蒙蒙
(北京市农林科学院蔬菜研究所,农业农村部华北地区园艺作物生物学与种质创制重点实验室,蔬菜种质改良北京市重点实验室,北京 100097)
基因编辑是一种可以在基因组水平上对DNA序列进行改造的遗传操作技术。目前,基因编辑技术已经在植物的遗传基础理论研究和生产实践中得到广泛应用。通过基因编辑技术进行基因功能研究和利用基因的靶向修饰进行性状改良是基因编辑应用最广泛的领域,在模式植物和大田作物中都有大量报道。蔬菜是人们获取人体所需要的维生素和微量元素的主要来源,基因编辑技术目前已成功应用在多种蔬菜作物中。随着功能基因组研究日渐深入,通过基因编辑技术进行功能基因研究和性状改良是蔬菜分子育种的必然方向。
1 基因编辑技术的发展历程
基因编辑的原理就是构建一个人工核酸内切酶,在预定的基因组位置切断DNA,产生DNA 双链断裂(double-strand break,DSB),DSB 在被细胞内的DNA 修复系统修复过程中会产生突变,从而达到定点改造基因组的目的。基因编辑技术应用的关键是核酸内切酶的使用,常用的核酸内切酶能够有效切割DNA,但它们通常在多个位点进行识别和切割,特异性较差。基因编辑的发展历程就是研究者对不同类型核酸内切酶的发现和改造的过程。基因编辑常用的核酸内切酶主要分为四类:巨型核酸酶(meganuclease,MegNs)(Choulika et al.,1995)、锌指核酸酶(zinc finger nucleases,ZFNs)(Wah et al.,1998)、转录激活因子样效应物核酸酶(transcription activator-like effector nucleases,TALENs)(Zhang et al.,2014)和成簇规律性间隔短回文重复系统(clusters of regularly spaced short palindromic repetition systems,CRISPR/Cas)(Cho et al.,2013)。
1.1 巨型核酸酶
在20 世纪80 年代后期发现的巨型核酸酶是一种脱氧核糖核酸内切酶,可以识别一段12~40 bp的DNA 序列,这种专一的长序列在DNA 链上重复出现的频率低,保证了其切割的特异性,但由于其对特定长序列的需求和巨大的酶体,导致其剪切效率低,应用难(Silva et al.,2011)。
1.2 锌指核酸酶
由于巨型核酸酶的缺陷,锌指核酸酶取代了巨型核酸酶。ZFNs 是一种人工融合蛋白,包括DNA识别域和核酸内切酶I。DNA 识别域由一系列Cys-His 锌指蛋白(zinc-fingers,ZFPs)串联组成,每个ZFP 识别并结合一个特异的三联体碱基。Kim等(1996)首次将3 个串联的锌指结构域(能够识别9 个核苷酸)与I 的C 端内切酶结构域通过一段连接蛋白融合,制造出第一个嵌合型核酸内切酶—ZFNs,并在体外证明该酶对靶DNA 具特异剪切能力。ZFNs 的DNA 识别域能识别特异位点并与之结合,而由I 构成的切割域具有剪切功能,针对目的基因设计特异性ZFPs,再将DNA 识别域与I 结合就可使靶位点产生DSB。细胞可以通过同源重组(HR)修复机制和非同源末端连接(NHEJ)修复机制来修复DNA。HR 修复有可能会对靶位点进行恢复修饰或者插入修复,而NHEJ修复极易发生插入突变或缺失突变,从而达到基因改造的目的。
基于ZFNs 的基因编辑技术可应用于很多物种及基因位点,具有较好的发展潜力。但是目前有3方面的缺陷制约了该技术的推广:一是,以现有的策略设计高亲和性的ZFNs,费时费力;二是,持续表达ZFNs 对细胞有毒性;三是,虽然三联体设计具有一定特异性,但仍然存在不同程度的脱靶效应(李想 等,2017)。
1.3 转录激活因子样效应物核酸酶
鉴于ZFN 技术的局限性,人们致力于寻找更为有效、特异性更强的基因编辑工具。2009 年,两组研究人员同时报道了TALE 蛋白能特异性识别并结合DNA 序列(Boch et al.,2009;Moscou &Bogdanove,2009),TALEN 技术由此应运而生。Christian 等(2010)将TALE蛋白与I核酸内切酶融合构成人工TALEN 核酸酶,一系列TALEN 蛋白串联起来可组成DNA 识别域,每一个TALEN 蛋白能识别并结合一段14~20 bp 的碱基序列,I 核酸内切酶形成二聚体时可以切割双链DNA。TALEN 技术是一种理论上能够对任意DNA目的序列进行靶向遗传修饰的基因编辑技术,是实现基因敲除、基因敲入或转录激活等靶向基因组编辑的里程碑。
目前TALEN 技术已经成功应用于酵母、哺乳动物和植物的位点特异性基因打靶,与锌指核酸酶系统相比有较大的应用优势,设计更简单、特异性更高,但仍然有脱靶效应、TALEN 与靶序列的特异性结合受到染色体位置及邻近序列影响等问题存在(李想 等,2017)。
1.4 成簇规律性间隔短回文重复系统
CRISPR/Cas 系统是继ZFN 和TANEN 技术之后最新发现的第三代基因编辑技术,更简单有效,应用前景更广阔。
CRISPR 是细菌和古细菌基因末端的多组DNA序列与反向序列以及30 bp 左右的空格序列(spacer DNA)串联成簇的、间隔规律的短回文重复序列。该结构最早是在Ishion 等(1987)对大肠杆菌的碱性磷酸酶基因研究中被发现。随后,Jansen等(2002)进一步研究了原核生物中的该重复DNA 序列,将该结构命名为CRISPR,同时确定了4 个CRISPR相关基因(-,)。在不同物种中根据Cas 蛋白不同将CRISPR 系统分为3 大类、10 小类,其中依赖Cas9 蛋白的CRISPR 系统属于第2 类,最简单常用(Lino et al.,2018)。CRISPR/Cas9 系统由Cas9 核酸内切酶、具有靶向特异性的CRISPR RNA(crRNA)和反式激活CRISPR RNA(trans-activating crRNA,tracrRNA)3 种 元件组成。crRNA 通过碱基配对与tracrRNA 结合形成tracrRNA/crRNA 复合物,通过人工设计这两种RNA,可以将其改造、整合形成具有引导作用的sgRNA(single guide RNA),能引导Cas9 蛋白对DNA 的定点切割(Jinek et al.,2012)。sgRNA的特异识别功能依赖靶序列下游3′末端的一段保守结构—PAM(proto-spacer adjacent motifs),tracrRNA/crRNA 复合物在PAM 序列上游切割DNA造成DSB,并启动DNA 损伤修复机制。
CRISPR/Cas9 系统设计简单,在多基因编辑应用方面更简单高效,仅需设计sgRNA,就能同时进行多基因编辑,并且靶向精确、脱靶率低、细胞毒性低、更廉价简便,深受研究人员的青睐。
2 基因编辑技术在蔬菜中的研究应用
目前,基因编辑技术已经在植物的遗传基础理论研究和生产实践中广泛应用,植物基因的靶向修饰是基因编辑技术应用最广泛的领域。基因编辑技术可以通过修饰内源基因来帮助设计所需的植物性状,在模式植物和大田作物中都有大量报道;蔬菜作物种类繁多,基础研究较大田作物稍落后,主要在茄科、十字花科及葫芦科这三大科常见蔬菜中研究较多。本文对相关文献进行汇总,介绍了基因编辑技术在以上三大科主要蔬菜作物中的研究概况(表1)。
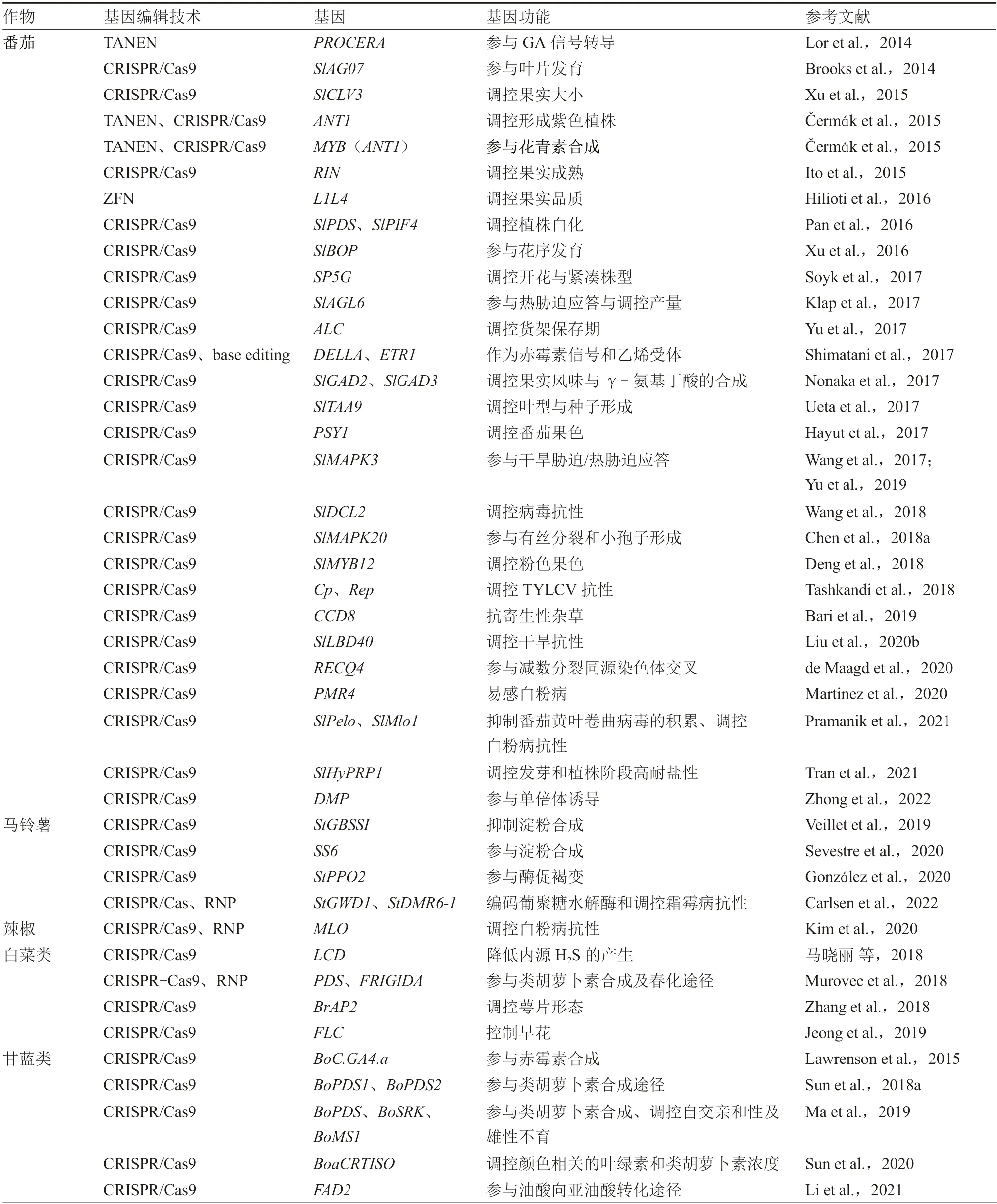
表1 主要蔬菜作物中的基因编辑研究

续表
2.1 茄科蔬菜中基因编辑技术的研究应用
茄科蔬菜的基因编辑技术主要在番茄、马铃薯、辣椒上报道较多。
2.1.1 番茄中基因编辑技术的研究 番茄()属于茄科番茄属二倍体作物,在全世界广泛栽培,其基因组已经完成测序,遗传转化方法也比较成熟,是理想的蔬菜基因编辑技术研究材料。目前在番茄中已有大量基因编辑相关文献报道(张爱萍 等,2022)。
Hilioti 等(2016)首次证实了基于ZFNs 的基因编辑技术在番茄上的有效性,通过对番茄--()基因不同位点的编辑改造,证实了的DNA 结合域上游序列的改变可导致包括番茄果实在内的多种表型变异。
Lor 等(2014)首次在番茄基因编辑中成功使用TALEN 技术,该研究的靶基因()是番茄中唯一报道的DELLA 家族基因,而DELLA 蛋白是赤霉素(GA)信号的负调控因子。该研究通过基因编辑产生错义突变导致部分活性丧失,从而确定在番茄GA 信号转导中的作用。
Brooks 等(2014)首次利用靶向基因(),将CRISPR/Cas9技术应用于番茄基因编辑,发现敲除的番茄突变体植株叶片呈针状而不是正常的扁平复叶。Čermák等(2015)比较了TALENs 和CRISPR/Cas9 在番茄上的基因编辑效率,该研究在控制花青素生物合成的基因上游插入一个强启动子,导致花青素在番茄组织中过表达和异位积累。在这次基因编辑中,超过2/3 的插入是精确的,并且没有检测到脱靶修饰,TALENs 和CRISPR/Cas9 都以相似的效率实现了基因靶向编辑。
随着基因编辑技术的发展,CRISPR/Cas9技术逐渐取代了ZFN和TALEN技术。Danilo 等(2018)以番茄花色苷合成基因为靶基因,利用CRISPR/Cas9 系统在番茄上实现了同源介导(homology directed repair,HDR)的外源基因插入。Li 等(2018a)通过CRISPR/Cas9 系统在番茄中进行多基因编辑,该研究同时敲除了类胡萝卜素代谢途径中的多个基因,抑制了番茄红素向α-胡萝卜素和β-胡萝卜素的转化,促进了番茄红素的生物合成和积累。随后,以CRISPR/Cas9 为主的基因编辑技术就在番茄上广泛应用,在产量、抗病性、非生物胁迫等基因挖掘到性状改良上都取得了快速进展(Shimatani et al.,2019;Pramanik et al.,2021;Tran et al.,2021)。最近,根据玉米和拟南芥上的研究基础,Zhong 等(2022)通过CRISPR/Cas9 基因编辑技术改造()基因,首次在番茄上创制了单倍体诱导系,揭开了蔬菜倍性育种的新篇章。
2.1.2 马铃薯中基因编辑技术的研究 马铃薯(L.)是世界上重要的主食作物之一,属于茄科茄属的杂合四倍体作物。目前利用CRISPR/Cas9 技术已经成功在马铃薯中实现了定向诱变。
Veillet 等(2019)采用CRISPR/Cas9 技术成功编辑了马铃薯的颗粒结合型淀粉合成酶(granulebound starch synthase,GBSS)基因,使淀粉合成受阻。Sevestre 等(2020)利用CRISPR/Cas9 技术敲除了1 个新发现的马铃薯淀粉合成酶基因,其结果显示:在基因第二外显子末端有一个保守序列(GT1),该序列作为靶点在CRISPER/Cas9 介导的基因编辑中表现出较高的基因编辑效率。
多酚氧化酶类(polyphenol oxidases,PPOs)是一种含铜酶,广泛分布于高等植物中,能够催化多种酚类化合物氧化成各自的醌类物质。醌类物质可以进行自聚合,与蛋白质中的氨基酸或自由基反应,形成深色沉淀物,这一过程即为“酶促褐变”,是水果和蔬菜品质降低的原因。González 等(2020)通过CRISPR/Cas9 系统敲除基因,显著降低了马铃薯块茎的PPOs 活性,褐变程度也明显减轻。
马铃薯是一种高度杂合的四倍体作物,其品种内和品种间基因组呈现高度杂合性和多态性,也是基因编辑策略和设计中要重点考虑的技术难点。Carlsen 等(2022)发现在原生质体这类细胞池水平上的高编辑效率,对在四倍体中实现全等位基因敲除至关重要,该研究通过聚乙二醇(PEG)介导、CRISPR/Cas 核糖核蛋白颗粒(ribonucleoprotein complex,RNP)瞬时转化原生质体,获得编码葡聚糖水二激酶(GWD)和霜霉病抗性6(-)基因的编辑植株。高编辑效率减少了下游繁琐而复杂的离体再生程序,而且用两个或多个RNP 同时靶向两个靶区中的任意一个,均对基因编辑产生明显的正协同效应。这些初步发现可能会引发更大规模的研究,以促进和优化植物的精确育种。
2.1.3 辣椒中基因编辑技术的研究 辣椒(L.)属于茄科辣椒属,是亚热带和温带国家最广泛栽培的茄科物种。辣椒于2014年完成基因组测序,为其基因编辑技术研究奠定了基础。
Li 等(2020)基于辣椒Zunla-1 参考基因组完成了辣椒全基因组的编辑位点鉴定和特异性分析,确定了辣椒基因组的1 组高度特异的CRISPR/Cas9编辑位点。该套编辑位点具有广泛的适用性,有助于将辣椒中CRISPR/Cas9 的脱靶率降至最低,有效应对CRISPR/Cas9 系统的脱靶效应。
辣椒的遗传转化受组织培养条件的限制,不同基因型组织培养能力差距大。Woo 等(2015)发现Cas9/sgRNA 核糖核蛋白复合物RNP 直接应用于原生质体转化可以获得无外源基因插入的编辑植株,减少了前期组织培养步骤,也是辣椒基因编辑技术研究的适宜方向。Kim 等(2020)利用来源于叶片和愈伤组织的辣椒原生质体做受体,进行RNP 转染,在辣椒CM334 和甜椒Dempsey 中实现了对植物病原尤其是白粉病病原具有广谱抗性的目标基因的编辑。该研究为在一定程度上突破组织培养的限制、实现RNP 途径的基因编辑技术在辣椒上的应用奠定了基础。
尽管目前已有辣椒全基因组序列和基因组编辑工具,但由于缺乏稳定的辣椒转化体系,致使辣椒的精确基因编辑仍处于起步阶段。为此,Park 等(2021)采用3 种农杆菌菌株AGL1、EHA101、GV3101,选择2 个辣椒品种辣椒CM334、甜椒Dempsey,选用pBAtC 作为CRISPR/Cas9 基因编辑载体,优化了筛选剂磷化氢蓖麻毒素(PPT)浓度,结果显示:GV3101 在甜椒Dempsey 中得到了更好的转化编辑效果。
2.2 十字花科蔬菜中基因编辑技术的研究应用
十字花科蔬菜种植最为广泛的两个属分别是芸薹属()和萝卜属(),其中萝卜属的基因编辑研究鲜有报道。芸薹属包括多种重要蔬菜作物,主要有3 个二倍体基本种,即白菜(,2=20,AA)、甘蓝(,=18,CC)、黑芥(,2=16,BB),以及三者之间相互杂交、经过不断选择进化得到的3 个异源四倍体复合种,即甘蓝型油菜(,2=38,AACC)、芥菜型油菜(,2=36,AABB)和埃俄比亚芥(,2=34,BBCC)。目前基因编辑研究报道较多的是在白菜、甘蓝和甘蓝型油菜上。
2.2.1 白菜类蔬菜中基因编辑技术的研究 白菜类蔬菜包括7 个变种,常见的有大白菜(L.ssp.)、普通白菜(L.ssp.Makino)、菜薹(L.var.Tsen et Lee)等。近年来,随着基因编辑技术的发展,白菜基因组和育种研究都取得了快速的发展。
大白菜作为一种重要的大宗蔬菜,种植范围广泛、历史悠久。生产者最初选择了开花晚的大白菜品种来提高产量,但是晚开花的大白菜需要经过冷处理数周才能诱导开花,最终收获种子。这种晚熟春化诱导开花的特性延长了大白菜的育种年限,阻碍了大白菜相关遗传研究的快速开展。为了解决这一问题,Jeong 等(2019)利用CRISPR/Cas9 系统将目标突变引入()基因,培育出不需要春化即可早开花的大白菜(和的双敲除突变体),加快了繁育进程。同时,该研究还发现和之间存在功能冗余。
硫化氢(hydrogen sulfide,HS)是植物细胞内源信号分子,对植物的生长、发育和抗逆性具有重要的调节作用,而L-半胱氨酸脱巯基酶(L-cysteinedesulfydrase,LCD)是合成内源HS的关键酶(尚玉婷 等,2018)。马晓丽等(2018)采用CRISPR/Cas9 基因编辑技术对大白菜基因进行敲除,基因编辑植株中内源HS 含量有不同程度的降低,为研究HS 信号在大白菜中的生理作用及育种应用提供了理论基础。
姜明亮(2020)选取孤基因(orphan genes)()及其高度同源基因()为靶基因,在大白菜中创制高效靶向突变,降低了果糖、葡萄糖和可溶性糖的含量,但蔗糖含量和蔗糖合成酶活性明显增加,推测可能依赖蔗糖合成途径影响可溶性糖代谢。该研究为白菜功能基因组的研究提供了重要技术支撑,同时对深入研究孤基因影响可溶性糖代谢的作用机制提供了材料和理论参考。
菜薹又称菜心,为不结球白菜类型中以幼嫩花茎为主要食用器官的常规栽培种,虽然在食用广度和栽培面积上不如大白菜,但其生育周期短,播种至产品收获仅需40~60 d,是研究白菜类蔬菜转化技术的理想材料。国家蔬菜工程技术研究中心生物技术室在菜薹原位转化成功的基础上(Liu et al.,1998),结合CRISPR/Cas9 系统,以四九菜心为试材,番茄红素脱氢酶基因()为靶基因,采用真空渗透原位转化方法,获得了无外源片段插入的基因编辑植株(宗梅 等,2022)。
2.2.2 甘蓝类蔬菜中基因编辑技术的研究 甘蓝类蔬菜为十字花科芸薹属的一年生或两年生草本植物,包括多个变种,常见的有结球甘蓝(var.L.)、羽衣甘蓝(var.DC.)、花椰菜(.var.L.)和青花菜(L.var.Plenck)等。甘蓝自交不亲和性强,必须通过低温诱导开花,生育期长,变种内遗传基础狭窄,基因编辑技术对高效创制甘蓝的遗传突变、实现优良品种改良起到重要作用。
Ma 等(2019)设计了1 种基于内源tRNA 处理的CRISPR/Cas9 基因编辑系统,实现了对结球甘蓝的多位点和多基因突变。该研究以番茄红素脱氢酶基因、S-位点受体激酶基因和雄性不育相关基因为靶基因,采用串联tRNA-sgRNA 结构的载体表达多个sgRNA,结果显示:基因敲除的植株表现白化矮小,作为可视标记直接验证该基因编辑系统的可行性;基因突变完全抑制了结球甘蓝的自交不亲和性,将自交不亲和系转化为自交亲和系;而基因突变产生了1 个雄性不育突变体,由于同时发生基因突变,该突变体在开花期与其非突变等位基因杂交兼容,从而使该雄性不育系能够通过蜜蜂介导的异花授粉进行繁育。
Sun 等(2018a)首次报道了CRISPR/Cas9 技术在羽衣甘蓝上的应用,该研究以类胡萝卜素合成相关的基因为靶点,使羽衣甘蓝两个同源基因同时发生定点突变,为羽衣甘蓝基因功能研究和分子育种提供了技术支撑。该课题组在此技术基础上,对羽衣甘蓝类胡萝卜素异构酶基因()进行编辑,共获得13个突变体,突变率高达81.25%;双等位基因突变株和纯合突变株的总类胡萝卜素和叶绿素含量均降低,总水平下降了11.89%~36.33%,叶片颜色从绿色变为黄色,创制了黄色羽衣甘蓝新种质(Sun et al.,2020)。
单碱基编辑技术(base editing)是在CRISPR/Cas9 系统基础上添加一些酶,实现对DNA 的单个碱基进行替换,具有高效而又精确的基因编辑能力,可在动植物细胞内引入点突变,用于培育具有理想表型的基因编辑动植物。例如:Cas9 蛋白与胞嘧啶脱氨酶组成融合蛋白可以实现胞嘧啶(C)到胸腺嘧啶(T)的单碱基转换,被称为BE3(APOBEC1-XTEN-nCas9-UGI,第三代碱基编辑器)系统(Tian et al.,2018);腺嘌呤单碱基编辑器(简称ABE),可以对目标位点实现高效腺嘌呤(A)到鸟嘌呤(G)的单碱基转换(Liu et al.,2022)。北京市农林科学院蔬菜研究所生物技术研究室在花椰菜基因编辑技术体系的基础上,通过BE3 系统对花椰菜的乙酰乳酸合成酶(acetolactate synthase)基因和着丝粒特异组蛋白变体(centromerespecific histone H3 variant)基因进行单碱基编辑突变,突变率为22%~87%,创制了高抗除草剂的花椰菜新种质(Wang et al.,2022)。
2.2.3 甘蓝型油菜中基因编辑技术的研究 甘蓝型油菜(L.)作为重要的油料作物,基因组和基因编辑相关研究较多,在产量、品质、抗性、花色、育性等多种性状中都有研究报道(杨文文等,2021)。通过CRISPR/Cas9 系统,Karunarathna等(2020)同时敲除甘蓝型油菜中含油量相关的和两个基因,获得了种子含油量明显增加的突变体植株;Khan 等(2021)敲除CYP78A6 家族基因,突变体植株角果和种子变小,但单株种子数量和质量增加;Sriboon等(2020)敲除开花调控基因的5 个拷贝,发现敲除其中1 个拷贝()后,植株出现早花表型,而敲除其他拷贝,则对开花时间没有明显影响;Liu 等(2020a)发现,敲除甘蓝型油菜中两个参与类胡萝卜素合成的玉米黄质环氧化酶基因和,改变了花瓣中类胡萝卜素的积累,紫黄素含量降低,叶黄素含量升高,从而导致了橘色花的产生;Xin 等(2020)发现,敲除Y127 株系中的Ms5 基因或敲除Westar 株系中的Ms5 基因,均会导致雄性不育,进一步研究显示,Ms5 通过与Ms5 或Ms5 形成无功能的异源二聚体而显性抑制Ms5 或Ms5 的功能,从而导致雄性不育。
Wu 等(2020a)利用基于鼠源胞嘧啶脱氨酶的单碱基编辑器BE3,将基因197 位密码子上的C 转化为T,获得的P197S 油菜突变体表现出耐除草剂苯磺隆的表型。Cheng 等(2021)利用基于人源胞嘧啶脱氨酶的单碱基编辑器,突变油菜基因,发现和的P197F(根据拟南芥的ALS 序列编号)突变均能赋予油菜抗苯磺隆除草剂的能力。且这两个拷贝同时突变的油菜植株比突变单个拷贝的植株能够耐受更高浓度的除草剂,说明突变介导的除草剂抗性具有一定的剂量效应。
2.3 葫芦科蔬菜中基因编辑技术的研究应用
葫芦科是世界上重要的可食用植物科之一,其重要性仅次于禾本科、豆科和茄科,其中包括黄瓜、南瓜、丝瓜、西瓜等常见的蔬菜和瓜果。然而,这些作物中大多数尚未建立高效的遗传转化体系。葫芦科蔬菜中基因编辑的研究报道大多在黄瓜和西瓜上。
2.3.1 黄瓜中基因编辑技术的研究 黄瓜(L.)属于葫芦科黄瓜属的二倍体蔬菜作物,其基因编辑的相关报道大都是利用CRISPR/Cas9技术。
以色列科学家采用CRISPR/Cas9 技术首次在黄瓜中开展抗病毒研究(Chandrasekaran et al.,2016)。该试验设计了两个sgRNA,以黄瓜(真核翻译起始因子4E)基因为靶基因,获得了一系列靶点缺失和插入突变的植株。用黄瓜叶脉黄化病毒(CVYV)、小西葫芦黄花叶病毒(ZYMV)和番木瓜环斑型花叶病毒(PRSV-W)接种突变株和野生型黄瓜植株,基因编辑纯合突变株表现出广泛的病毒抗性。
Hu 等(2017)利用CRISPR/Cas9 系统编辑了抑制黄瓜心皮发育的基因,成功获得了全雌系材料,提高了产量。短瓜基因编码1 种葫芦科特异的环型E3 连接酶,其突变可导致自身泛素化和降解增强,同时造成乙烯合成速率限制酶ACS2 的积累,Xin 等(2019)成功通过CRISPR/Cas9 技术获得了和这2 个基因的编辑突变体,并研究了其调控乙烯合成和黄瓜果实伸长的分子机制。Zhang 等(2020a)成功实现了对短瓜基因的CRISPR/Cas9 编辑,验证了通过促进细胞增殖调控黄瓜果实伸长。基因属于TCP(PROLIFERATING CELL FACTORS)基因家族,Yang 等(2020)成功获得基因的CRISPR/Cas9 突变体,揭示了基因通过直接调控乙烯的合成来控制黄瓜茎卷须的形态和攀缘。
毛状根诱导系统因其生长快、遗传稳定性高,可作为研究基因表达和功能的有效方法,也被证实是评估CRISPR/Cas9 系统用于基因组编辑活动的有效工具(Jacobs &Martin,2016)。Nguyen 等(2022)对黄瓜根瘤菌介导的毛状根转化系统进行了优化,为验证植物转化载体、CRISPR/Cas9 构建活性以及选择黄瓜基因编辑的靶向gRNA 提供了有效工具,建立的毛状根转化系统为黄瓜和其他植物基因组的编辑技术优化和进一步研究提供了重要信息和新的思路。
2.3.2 西瓜中基因编辑技术的研究 西瓜()属于葫芦科西瓜属一年生蔓生藤本植物。西瓜是葫芦科中第2 个基因组编辑成功的物种(Tian et al.,2017,2018)。
Tian 等(2017)采用CRISPR/Cas9 技术对西瓜的(phytoene desaturase)基因进行精确编辑。该研究获得的转化苗几乎达到了100%的编辑效率,所有的转基因西瓜植株都带有S 突变,并表现出明显的或镶嵌式的白化表型。随后,Tian等(2018)利用基于CRISPR/Cas9 的单碱基编辑器特异性突变乙酰乳酸合酶(acetolactate synthase,ALS)基因的Pro190 位点,获得了抗除草剂苯磺隆的西瓜新种质。Zhang 等(2020c)采用CRISPR/Cas9 系统敲除了编码植物磺肽素(Phytosulfokine,PSK)前体的基因,以增强西瓜对尖孢镰刀菌(,FON)的抗性。
虽然西瓜遗传转化取得了一些进展,但从组织培养到再生仍然具有挑战性。Feng 等(2021)根据小麦中的研究结果,通过共表达生长调节因子4(GRF4)和GRF-相互作用因子1(GIF1),改善西瓜再生,在西瓜中实现了高效转化,并与基因组编辑工具相结合,在西瓜中实现了高效的基因编辑,成功地创制了二倍体无籽西瓜。该研究为今后西瓜遗传转化的研究和育种提供了有力的理论基础。
3 总结与展望
基因编辑技术作为生命科学的颠覆性技术,已在农业领域得到了广泛应用。CRISPR-Cas 系统的出现加速了基因编辑技术的发展,基因编辑工具的开发和应用日新月异,具有更广泛的目标、更高的效率和特异性,以及更高的精度。目前,基因编辑技术已被应用于植物中各种可遗传的基因组修饰,例如:随机插入/缺失(InDels),点突变或核苷酸替换,同源基因和多基因突变,碱基编辑器(base editor),先导编辑器(prime editor),HDR 介导的基因替换、点突变、DNA 插入和删除,以及有针对性的染色体重排等(Liu et al.,2022)。在技术的持续拓展及延伸下,基因编辑将在更广泛的领域带来更大的社会价值与经济价值。蔬菜作为人们日常饮食中必不可少的食物之一,是提供人体必需的多种维生素和矿物质的主要来源。蔬菜中的VK、VC、有机硫化物和硝酸盐对慢性疾病的预防有益,钾、镁、膳食纤维、类胡萝卜素等成分有利于预防心脑血管疾病。针对蔬菜的这些特征定向育种,对一些优良性状进行聚合和改良,提高蔬菜的产量、抗性、品质和口感一直是育种家和消费者共同追求的目标。传统育种方法不但易受种间生殖隔离的限制、不良基因连锁的影响,而且育种年限很长。基因编辑技术优势明显,可以克服以上困难,在性状改良上表现出良好的靶向性和易用性,发展空间巨大,应用前景广阔。
蔬菜作物种类繁多,基因组学研究基础差距大,较大田作物在遗传转化和基因编辑研究上都相对滞后。但基因编辑工具是相通的,最新的基因编辑工具已经很快应用到蔬菜作物中,如花椰菜和西瓜中的单碱基编辑器(Tian et al.,2018;Wang et al.,2022),番茄和甘蓝型油菜上的同源基因和多基因编辑(Zhai et al.,2020;Tran et al.,2021),同源介导基因编辑的外源基因插入(Danilo et al.,2018),以及RNP的应用(Kim et al.,2020),等等。虽然蔬菜基因编辑研究上取得了很多亮点,但在一些蔬菜中还存在缺乏高效遗传转化技术的瓶颈问题,如辣椒、西瓜、南瓜、白菜、茄子等,缺乏适用于多种基因型的高效遗传转化体系,同时可用于编辑的功能基因研究也是核心问题。
围绕当前基因编辑技术在蔬菜作物中存在的问题,无转化体系的蔬菜作物应尽快建立适宜的转化体系。首先可以根据不同蔬菜作物的特性,研究建立不同外植体、不同转化途径的技术体系。例如:番茄以子叶、叶片和下胚轴为外植体,花椰菜以下胚轴和花序轴为外植体都有成功的转化体系建立(Brooks et al.,2014;Hilioti et al.,2016;Wang et al.,2022)。对一些难以转化和再生的作物,如辣椒、茄子、西葫芦等,可探索以大孢子、小孢子、原生质体、花粉粒等为受体的转化编辑体系。同时不依赖遗传转化、直接获得无外源基因插入的基因编辑材料显得更具优势,例如:通过农杆菌介导的基因的瞬时表达(Iaffaldano et al.,2016;Chen et al.,2018b),以及RNP 原生质体侵染等(Woo et al.,2015;Murovec et al.,2018)。本课题组也在芸薹属菜薹上探索不依赖组织培养的原位转化编辑体系,并取得了一些进展(宗梅 等,2022)。因此,建立和完善不同蔬菜作物的遗传转化技术,努力探索不依赖于组织培养的转基因途径是蔬菜作物基因编辑技术广泛应用的研究方向。
另外,随着基因组的大规模测序,蔬菜功能基因组的研究也迅速发展。通过基因组测序和大数据分析,根据大田和模式作物的研究基础快速高效鉴定蔬菜中对应的功能基因,针对蔬菜作物性状特点结合正向遗传和反向遗传明确农艺性状的控制基因,再结合基因编辑技术鉴定基因功能和作用机制。可编辑的功能基因研究和基因编辑技术的发展应用是相互促进、相辅相成的。
虽然基因编辑技术在蔬菜上的研究已经取得了大量进展,但在不同种类蔬菜作物上的广泛应用还有很多问题需要解决。随着蔬菜基因组研究的逐渐深入和发展,技术的不断完善和延续,基因编辑技术作为最具潜力的基因组改造工具必将在蔬菜作物功能基因研究和分子育种应用中发挥重大作用。
