大田野生麦苗穗发芽抗性的鉴定与遗传分析
2022-08-19张希太谢淑芹蔺桂芬
张希太,肖 磊,董 策,谢淑芹,蔺桂芬
(邯郸市农业科学院生物技术研究室,河北邯郸 056001)
0 引言
小麦成熟期穗发芽(pre-harvest sprouting,PHS)是一种世界性的气候灾害。联合国粮农组织统计,每年全球因穗发芽造成的损失占世界小麦总产量的20%,约相当于中国小麦年产量[1]。中国有85%的小麦面积都存在穗发芽隐患[2],黄淮麦区、长江中下游麦区曾多次大面积发生小麦穗发芽灾害,西南冬麦区及东北春麦区穗发芽灾害也频繁发生[3]。小麦穗发芽后不仅产量降低,更主要是造成品质严重变劣,甚至丧失食用、加工利用的价值。穗发芽灾害不发则已,一发则损失惨重。穗发芽灾害发生的外因是连续阴雨和潮湿的气候,内因是小麦品种本身对穗发芽的抗性差。当今科技影响气候的能力有限,因此,只能通过抗穗发芽育种提高小麦品种自身的穗发芽抗性来抵抗灾害的发生。小麦抗穗发芽育种存在着2 个技术瓶颈,首先是用于育种的高抗穗发芽种质资源严重匮乏;其次是在非小麦穗发芽灾害发生年份,育种后代材料抗穗发芽性状的大田定向选择困难。高抗穗发芽种质资源的严重匮乏是限制小麦抗穗发芽育种未能取得突破性进展的主要障碍。因此创制或收集高抗穗发芽的小麦种质资源是当前小麦抗穗发芽育种的首要问题。
小麦抗穗发芽研究一直受到世界各国的重视。中国科技工作者在小麦抗穗发芽育种方面做了大量工作。首先对现有的大量小麦品种资源材料进行了抗穗发芽鉴定筛选。也选出了一些抗性相对较好的材料[3-8],但是受现有小麦遗传背景的限制并没有从根本上找到解决穗发芽问题的抗源基因。
2011 年,国家小麦工程技术研究中心,利用基因枪法获得了转反义硫氧还蛋白基因(anti-trxs)的抗穗发芽转基因小麦材料。通过人工模拟降雨穗发芽试验,转anti-trxs基因小麦种子比非转基因对照小麦种子的萌动时间明显延长,穗发芽受到明显的抑制[9-10]。2013年,美国农业部农业研究局和堪萨斯州立大学的研究人员发现并克隆出一个被命名为PHS 的能防止植物提前发芽的基因[11]。但是这些材料转入的是人工克隆的工程基因,对生物环境的安全性还需评估,特别是目前国家严格限制在粮食作物上转工程基因的形势下,该材料在小麦抗穗发芽育种上的应用还需要等待。
通过观察人们发现小麦的伴生杂草‘节节麦’具有极强的抗穗发芽特性。四川农业大学利用远缘杂交技术帅先将‘节节麦’的高抗穗发芽基因引入小麦,创制出了小麦高抗穗发芽材料RSP[12-15],为小麦抗穗发芽育种提供了宝贵的种质资源;河南大学在‘节节麦’抗穗发芽基因研究与利用方面也做了大量工作[16-18]。
在“一年两熟”种植模式的黄淮麦区,一般当年5—6 月小麦收获后种植大秋作物(以玉米为主),9 月底10 月初大秋作物收获后再播种冬小麦。秋收后耕地前,大田中往往生长有少量的刚刚出土不久的野生麦苗,这些野生麦苗是小麦收获时由于粗放操作遗落于大田中的麦粒、麦穗萌发长成。这些遗落于大田中的麦粒、麦穗在土壤中经过了夏季充沛的降雨直到9月底10月初秋收后才萌发出土,应该具有很强的休眠性和抗穗发芽能力,于是笔者收集了大量的野生麦苗,进行集中移栽,对其遗传稳定性、穗发芽抗性、遗传亲缘关系进行了研究。旨在获取具有极强穗发芽抗性的种质资源。
1 材料与方法
1.1 野生麦苗的收集移栽与遗传稳定性观察
1.1.1 野生麦苗的收集与集中移栽 每年9 月底10 月初,秋收后耕地前,对计划收集野生麦苗的农田(夏茬为冬小麦)进行普查,将发现的野生麦苗进行根部带土团移栽。大田中生长的野生麦苗分2 种类型:一类是单棵生长的野生麦苗(由落入大田的一粒种子萌发长成,图1B),第二类是成簇生长的野生麦苗(由落入土中的整个麦穗上的种子萌发生长而来,图1A)。第一类野生麦苗移栽入观察圃时,对单株进行编号,记录发现地块及移栽时间等;第二类将簇分成单株移栽入观察圃,编簇号和株号,记录发现地块及移栽时间等。本试验野生麦苗的收集地为河北省成安县商城镇赵横城村北农田。分别于Ⅰ:2014年9月30日。Ⅱ:2015年9月30 日。Ⅲ:2016 年9 月30 日。Ⅳ:2017 年9 月30日。分4批次收集大田野生麦苗集中移栽到本研究室试验田的观察圃中。

图1 秋收后冬小麦播种耕地前大田中出土的野生麦苗
1.1.2 集中移栽的野生麦苗生长一致性观察与种子收获 野生麦苗集中移栽入观察圃后,立即浇水保证移栽苗的成活,加强管理,使移栽成活后的野生麦苗生长茁壮。从第一生育周期开始对第二类同一簇(穗)系内的野生麦苗单株进行一致性测试,参照中华人民共和国国家标准(GB/T 19557.2—2004)[19]进行。小麦成熟后,对于第一类野生麦苗,按株号进行单株收获并按株号保存种子。对于第二类野生麦苗,将生长一致、性状稳定的簇(穗)系内所有单株混收并按簇(穗)系号保存种子;对个别遗传性状不稳定、生长不一致的簇(穗)系进行淘汰或另行试验,本文不再阐述。
1.1.3 野生麦苗的遗传稳定性观察 第二个生育周期播种时,将第一类野生麦苗按株系播种;保留的第二类野生麦苗按不同的簇(穗)系播种,每系不播种少于2000株。整个生育期参照GB/T 19557.2—2004 标准观察两类野生麦苗各株系和簇(穗)系内植株性状的一致性,继续对个别遗传性状不稳定、生长不一致的株系与簇(穗)系进行淘汰。
第3个生育周期只对第一类野生麦苗株系内植株进行生长一致性观察。两类野生麦苗的后代,都需进行2年的生长一致性观察,如果连续2年株系或簇(穗)系内植株的遗传性状都表现一致,说明该株系或簇(穗)系已经遗传稳定。然后对遗传稳定的野生麦苗系进行穗发芽抗性鉴定。
1.2 遗传稳定野生麦苗系农艺性状调查与穗发芽抗性测定及种质系的选择
1.2.1 遗传稳定野生麦苗系主要农艺性状调查与相对穗发芽指数测定 将遗传稳定的野生麦苗系重新统一编号,按行距20 cm,株距10 cm单穴单粒播种,每系播种数量不少于500株。整个生育期调查株高、抗倒性、抗病性(主要调查白粉病、锈病、叶枯病、赤霉病4种病害)、越冬抗冻性、分蘖率、有效分蘖率、穗粒数、千粒重等主要农艺性状指标。
穗发芽抗性测定按照中华人民共和国农业行业标准(NY/T 1739—2009)[20]进行。计算见公式(1)和(2)。

1.2.2 野生麦苗抗穗发芽种质系的入选标准 本试验野生麦苗抗穗发芽种质系选择标准:[1]遗传稳定;[2]相对穗发芽指数<0.05;[3]株高≤75 cm;[4]抗倒性为1级;[5]大田无病害发生;[6]越冬抗冻性为1级;[7]分蘖率≥2000%;[8]有效分蘖率≥80%;[9]穗粒数≥35;[10]千粒重≥45 g。
1.3 野生麦苗抗穗发芽种质系的遗传背景(亲缘关系)分析
1.3.1 野生麦苗抗穗发芽种质系遗传的环境因素分析秋后萌发出土野生麦苗种子的长休眠特性,充分说明了在自然界小麦生产庞大的群体中存在着具有强种子休眠性的高抗穗发芽基因及表型。分析这些高抗穗发芽基因的来源:①可能是小麦本身基因发生的突变,但这种可能性几率很低;②小麦的伴生杂草野麦子(野燕麦Avena.fatua、节节麦Aegilops tauschiiCoss.等)的种子都具有极强的休眠特性和抗穗发芽特性[12,16-17]。这些与小麦属于同一亚科的抗穗发芽植物,在长期与小麦的共同生长中存在着天然杂交的可能,通过天然杂交将种子的强休眠抗穗发芽基因转给小麦的可能性较大。特别是节节麦和小麦分别属于小麦族的山羊草属与小麦属,亲缘关系较近,在小麦的进化史上,曾经成功的通过属间天然杂交为现代小麦贡献了D 染色体组,因此,节节麦和小麦发生天然杂交的可能性极大。通过观察大田中伴随小麦生长的野燕麦和节节麦发现,野燕麦的扬花期和小麦不同步(图2B)所以野燕麦和小麦发生天然杂交的可能性极小。野燕麦和小麦分别属于燕麦族和小麦族,亲缘关系较远,即使发生了远缘杂交,不通过人工幼胚抢救其杂种幼胚也不能发育成苗;而大田中生长的节节麦的扬花期和小麦同步(图2A),二者又有较近的亲缘关系和成功发生天然杂交的历史事实,因此现代发生天然杂交的可能性较大。四川农业大学的刘登才等[13]在进行节节麦和普通小麦的远缘杂交研究时,发现了有些节节麦类型和普通小麦远缘杂交的幼胚不需要胚抢救培养也能形成种子并发育成苗,因此大田中抗穗发芽的野生麦苗极有可能是混合生长的节节麦和普通小麦的天然杂交后代。

图2 大田中与小麦同时杨花的节节麦和野燕麦
1.3.2 野生麦苗抗穗发芽种质系的疑似亲本小麦品种的调查 调查收集地块在收集野生麦苗抗穗发芽种质系之前5~10 年间,种植过的小麦品种,这些种植过的小麦品种都有可能是和节节麦发生天然杂交的亲本材料。然后采用这些疑似亲本小麦品种的基因组DNA作参考进行遗传分析。
1.3.3 野生麦苗抗穗发芽种质系基因组DNA分子遗传分析
(1)SSR分子标记引物的选择
基于野生麦苗抗穗发芽种质系遗传的环境因素分析,在分子遗传水平上重点检测,野生麦苗抗穗发芽种质系和节节麦的遗传关系。因为节节麦和现代小麦具有同源的D染色体组,在发生天然杂交后,节节麦和小麦D染色体组间可以实现同源染色体配对,通过同源染色体间易位交换遗传物质的可能性极大。所以首先选用现有开发成功的小麦D 染色体组上的SSR 位点的引物进行PCR检测。
(2)检测材料DNA的提取与模板液制备
检测用的‘节节麦’和被检测野生麦苗抗穗发芽种质系都来源于同一地块。疑似亲本小麦品种从该品种育种者处收集。采用SDS 法并参照《植物基因工程原理与技术》[21]从暗培养的黄花苗中提取DNA,纯化后取少量的DNA 样品溶于无菌的超纯水中制成浓度约为20 ng/μL的模板液放于4℃冰箱中备用。
(3)SSR-PCR反应体系的构建与反应程序
PCR 反应体系的构建,采用25 μ L 反应体系,各成分的含量为:TaqDNA 聚合酶(5 U/μL) 0.2 μL、10×Buffer 2.5 μL、MgCl2(25 mmol/L) 2 μL、dNTP(2.5 mmoL) 1 μL、上下游引物(工作浓度)各1 μL、DNA 模板液(20 ng/μL) 0.5 μL、无菌超纯水16.8 μL。PCR 反应在英国TECHNE 公司的TC-5000 PCR 仪上进行,反应程序为:94℃预变性5 min,94℃变性50 s,退火温度(参考引物合成报告单推荐温度微作调整)退火时间为50 s,72℃延伸1 min,(30个循环),72℃后延伸5 min,4℃保存。
(4)SSR-PCR反应产物的电泳检测
PCR 反应产物采用8%聚丙烯酰胺凝胶进行电泳检测,每泳道上样量为8 μL,在200 V 恒电压下电泳5 h,剥胶,银染,照相,进行扩增条带的分析。
2 试验结果与分析
2.1 野生麦苗收集移栽与遗传稳定性统计
由表1数据知,当年的9月底10月初,在大田中生长的野生麦苗,根部带土移栽很容易成活,通过4年的移栽试验,不论是单棵生长还是成簇(穗)生长的野生麦苗,只要移栽后及时浇水成活率都为100%。
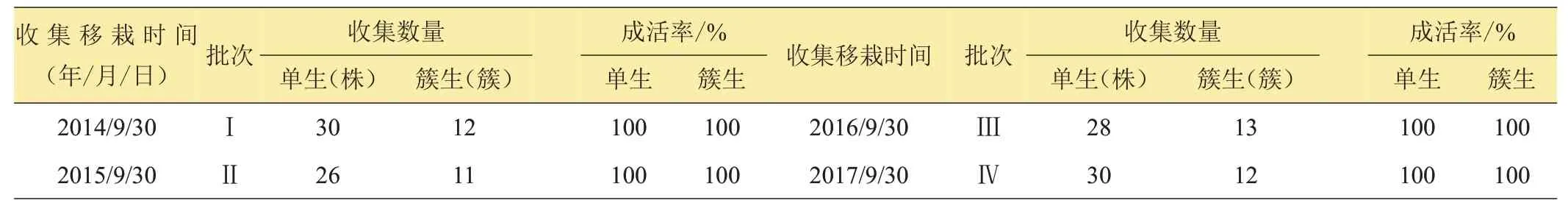
表1 野生麦苗收集移栽统计表
由表2数据知,大田中簇生野生麦苗的簇系后代,是由落入土中的一个麦穗上的种子生长而来,在进行第一年大田遗传稳定性观察时,90%以上簇系的后代都生长一致,不足10%簇系的后代中,出现个别性状表现不一致的单株,本试验中将后代生长不一致的簇系淘汰。第二年只对第一年未淘汰的簇系继续进行大田生长一致性观察。第一年后代生长一致的簇系,第二年其后代仍都生长一致。通过连续2年的大田生长一致性观察,大田中成簇生长的野生麦苗90%以上的簇系都生长一致且能稳定遗传。
大田中单棵生长的野生麦苗,移栽后的第一年由于没有形成后代群体,从第二年才开始进行后代生长一致性观察。由表2 数据知,第二年在首次进行后代生长一致性观察时,大田中单棵生长的野生麦苗只有63%以上的株系生长一致,本试验中将生长不一致的株系淘汰。第三年只对第二年首次观察未淘汰的株系进行第二次生长一致性观察。第二年后代生长一致的株系,第三年其后代仍然生长一致。通过连续2 年的大田生长一致性观察,大田中单棵生长的野生麦苗63%以上株系都能稳定遗传。
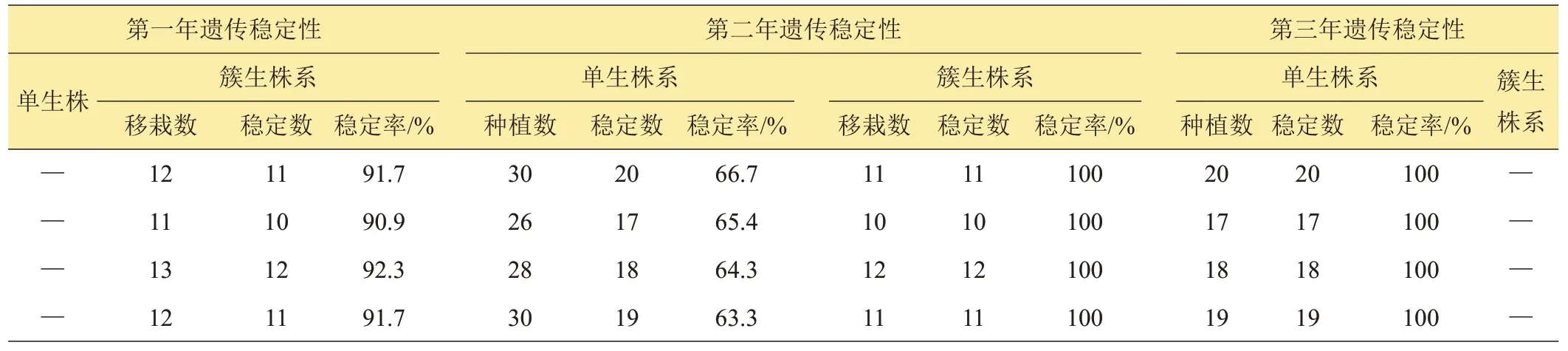
表2 野生麦苗遗传稳定性统计表
2.2 稳定遗传的野生麦苗系主要农艺性状调查及穗发芽抗性的测定结果
通过对生长一致、稳定遗传的118 个野生麦苗系主要农艺性状指标的调查,数据如表3。野生麦苗系的植株都比较高,有65.3%的野生麦苗系的株高都在100 cm 以上,有79.7%的野生麦苗系植株高于80 cm,20.4%的野生麦苗系株高低于80 cm,只有6.8%的野生麦苗系的植株不高于75 cm;野生麦苗系的抗倒性大部分较差,有65.3%的野生麦苗系发生中等以上倒伏,倒伏面积大于10%。21.2%的野生麦苗系倒伏较轻,只有13.6%的野生麦苗系不发生倒伏;野生麦苗系的抗病性(白粉病、锈病、叶枯病、赤霉病)普遍较好。在大田环境中,80.5%的野生麦苗系无病害发生,只有19.5%的野生麦苗系轻微发病;野生麦苗系的越冬抗冻性都较好,越冬冻害都不高于2 级,55.9%的野生麦苗系越冬冻害为1 级;野生麦苗系的分蘖率和有效分蘖率都较高,本试验所有野生麦苗系的分蘖都不低于20 倍,有效分蘖率都不低于80%;野生麦苗系的穗粒数较多,本试验所有的野生麦苗系穗粒数都不少于35粒,但野生麦苗系的千粒重较低,在行距20 cm,株距10 cm单穴单粒种植模式下,78.8%的野生麦苗系千粒重在45 g 以下,只有21.2%的野生麦苗系千粒重不低于45 g。

表3 稳定遗传的野生麦苗系主要农艺性状调查统计表
分析表4 中118 个野生麦苗系的穗发芽抗性测定结果,野生麦苗系的穗发芽抗性普遍较好。有19.5%的野生麦苗系的相对穗发芽指数为0,达到了特高抗水平。78.8%的野生麦苗系穗发芽抗性都能达到高抗水平,仅有17.8%和3.4%野生麦苗系的穗发芽抗性为抗和中抗水平。

表4 稳定遗传的野生麦苗系的穗发芽抗性测定结果统计表
2.3 野生麦苗抗穗发芽种质系的筛选结果
根据本试验制定的野生麦苗抗穗发芽种质系的选择标准,符合条件的野生麦苗抗穗发芽种质系应为以下集合的交集:{野生麦苗抗穗发芽种质系}={相对穗发芽指数<0.05的系}∩{株高≤75 cm的系}∩{抗倒性为1级的系}∩{无病害发生的系}∩{越冬抗冻性为1级的系}∩{分蘖率≥2000%的系}∩{有效分蘖率≥80%的系}∩{穗粒数≥35 的系}∩{千粒重≥45 g 的系}={21、46、63、106}。从后代生长一致、稳定遗传的118 个野生麦苗系中选出了‘ZZX21’、‘ZZX46’、‘ZZX63’、‘ZZX106’四个符合标准的野生麦苗抗穗发芽种质系,其抗穗发芽效果如图3所示。

图3 野生麦苗抗穗发芽种质系21、46、63、106抗穗发芽效果
2.4 野生麦苗抗穗发芽种质系收集地块以往种植情况调查
通过走访调查收集抗穗发芽种质资源系‘ZZX21’、‘ZZX46’、‘ZZX63’、‘ZZX106’地块的主人获悉,从收集之日起前5~10年间,本地块主要种植的小麦品种有‘济麦22’、‘邯6172’、‘衡4399’。
2.5 野生麦苗抗穗发芽种质系的SSR-PCR检测结果

表5 发现野生麦苗抗穗发芽种质系地块以往种植情况调查
分析引物Xgwm493(检测位点位于3B染色体)检查结果(图4A)、引物Xgwm538(检测位点位于4B 染色体)检查结果(图4B)、引物Xgdm33(检测位点位于1A、1D染色体)检查结果(图4 C),3对引物分别在4个抗穗发芽种质系‘ZZX21’、‘ZZX46’、‘ZZX63’、‘ZZX106’的基因组DNA中检测到了和‘节节麦’一致而参考亲本‘济麦22’、‘邯6172’、‘衡4399’没有的特异性条带。引物Xgwm493在‘节节麦’、4个抗穗发芽种质系的基因组DNA 中检测到了约160 bp 大小(图4A 中箭头所示)的特异性条带;引物Xgwm438 在‘节节麦’、4 个抗穗发芽种质系的基因组DNA 中检测到了约400 bp 大小(图4 B 中箭头所示)的特异性条带;引物Xgdm33 在‘节节麦’、4 个抗穗发芽种质系的基因组DNA中检测到了约800、720、80 bp大小的特异性条带(图4 C 中箭头所示);这充分证明了抗穗发芽种质系‘ZZX21’、‘ZZX46’、‘ZZX63’、‘ZZX106’和‘节节麦’的亲缘关系。
引物Xgdm33(检测位点位于1A、1D染色体)和引物Xgwm52(检测位点位于3D 染色体)在4 个抗穗发芽种质系的基因组DNA 中不仅检测到了和‘节节麦’一致而参考亲本‘济麦22’、‘邯6172’、‘衡4399’没有的特异性条带,还检测到了一些和全部或部分参考亲本一致而‘节节麦’没有的特异性条带(图4D、E 箭头所示);引物Xgwm314(检测位点位于3D染色体)检测到了大量的和参考亲本‘邯6172’、‘衡4399’一致而‘节节麦’没有的特异性条带(图4E 箭头所示)。这充分证明了4个抗穗发芽种质系和‘济麦22’、‘邯6172’、‘衡4399’的亲缘关系。

图4 SSR分子标记检查结果
引物Xgdm33、Xgwm52、Xgwm314 在4 个抗穗发芽种质系的基因组DNA中都检测到了和‘邯6172’的亲缘关系,因此,抗穗发芽种质系‘ZZX21’、‘ZZX46’、‘ZZX63’、‘ZZX106’是‘节节麦’和‘邯6172’天然杂交后代的可能性较大。
3 结论
本研究通过对大田野生麦苗集中移栽,遗传稳定性观察与选择,稳定系的穗发芽抗性鉴定,筛选出了4 个对穗发芽有绝对抗性的小麦种质资源系‘ZZX21’、‘ZZX46’、‘ZZX63’、‘ZZX106’,为小麦抗穗发芽育种提供了宝贵的种质资源,丰富了小麦抗穗发芽育种遗传的物质基础。
通过遗传分析与分子标记检测,从分子水平上初步证明了从大田野生麦苗中获得的小麦高抗穗发芽种质资源材料是当地大田中的野生‘节节麦’和种植的普通小麦品种进行天然远缘杂交的后代。
4 讨论
4.1 前人研究结果
具有绝对抗性的高抗穗发芽种质资源的匮乏是限制小麦抗穗发芽育种取得突破的主要障碍。获得具有绝对抗性的高抗穗发芽种质资源一直是小麦育种工作者努力的方向。首先对现有的大量小麦品种资源材料进行了抗穗发芽鉴定,以期筛选出抗穗发芽特性好的材料。中国农业科学院品种资源研究所、西北农林科技大学、西南大学生物科技学院、山西省小麦研究所等单位都在小麦抗穗发芽种质资源的筛选鉴定方面做了大量的工作[3-8]。发现了一些穗发芽抗性与种子萌发生理上的关系,抗性与麦穗形态、颖壳、种皮颜色上的关系,也选出了一些抗性相对较好的材料。但是受现有小麦遗传背景的限制并没有从根本上找到抗穗发芽的抗源基因。
随着生物技术的发展,基因工程技术应用于小麦抗穗发芽种质资源的创制。美国农业部农业研究局和堪萨斯州立大学通过对小麦全基因组测序研究克隆成功了PHS 基因,转PHS 小麦的穗发芽受到明显抑制;国家小麦工程技术研究中心克隆的反义硫氧还蛋白基因(anti-trxs)转入小麦后,获得的00T89、01TY18 转基因株系,穗发芽率较对照分别降低35.5%和62%[9-11]。基因工程技术拓宽了抗穗发芽育种遗传的物质基础,具有广阔的发展前景。出于转基因安全性的考虑,目前转基因技术在小麦这种主要粮食作物上的应用受到了国家的严格限制。
自然界中存在着和小麦同族不同属的节节麦(分属于小麦属与山羊草属)有着极强的高抗穗发芽特性[12,16-17]。利用节节麦的高抗穗发芽基因创造小麦高抗穗发芽种质资源,一直是中国科技工作者研究的热点。四川农业大学的兰秀锦[12]对节节麦的抗穗发芽基因进行了染色体定位研究,确定了节节麦的抗穗发芽基因是隐性单基因位于2D染色体上,以四倍体的地方小麦品种矮兰麦做母本与采自河南的节节麦进行杂交,F1代用秋水仙素进行染色体加倍获得了双二倍体小麦RSP其穗发芽率仅为2.86%[14-15];河南大学的刘新浩[17]对节节麦的抗穗发芽特性进行了鉴定及遗传多样性研究,发现了生长于黄河中游的节节麦抗穗发芽特性都较国外品种好。河南大学的陈趁趁[18]对节节麦和小麦杂交后代的抗穗发芽特性进行了鉴定并对抗穗发芽基因所在的相关染色体片段分析,通过连续两年对群体的穗发芽分析,筛选到20份高抗穗发芽的材料,5天发芽率均在10%以下,在5D和6D染色体上,存在着6 对与极抗穗发芽基因紧密连锁的SSR 分子标记位点。以上研究都在节节麦和小麦的远缘杂交后代中获得了高抗甚至于极抗穗发芽材料,丰富了小麦抗穗发芽育种的遗传背景,有效利用了节节麦中的极抗穗发芽基因,为小麦抗穗发芽育种提供了宝贵的种质资源。
4.2 本研究较前人研究的先进性
本试验研究了一种从自然存在的大田野生麦苗中收集、筛选小麦高抗穗发芽种质资源的方法;并从分子水平上证明了大田野生麦苗中获得的小麦高抗穗发芽种质资源材料是当地大田中的野生节节麦和大田种植的普通小麦品种进行天然远缘杂交的后代,具有和节节麦相同的极强抗穗发芽特性。本技术方法简单易行,成本低廉,有效地利用了节节麦中的远缘极抗穗发芽基因资源,选出的小麦高抗穗发芽种质资源的遗传背景丰富多样,为当前的小麦抗穗发芽育种提供了宝贵有绝对抗性的抗穗发芽种质资源。
