高丹草“土-草-畜”界面的氮素运移规律*
2022-08-13赵海明游永亮武瑞鑫刘贵波杨志敏祖晓伟
李 源, 赵海明, 游永亮, 武瑞鑫, 刘贵波**, 杨志敏, 祖晓伟
(1. 河北省农林科学院旱作农业研究所/河北省农作物抗旱研究重点实验室 衡水 053000; 2. 张家口市农业科学院 张家口075000; 3. 河北省畜牧总站 石家庄 050030)
氮素是植物生长发育所需的最重要的营养元素之一, 氮肥对作物产量的贡献高达30%~50%。以往在氮素利用效率研究上常采用差值法计算, 然而, 此方法难以区分作物吸收的氮是来自肥料氮还是土壤氮, 导致不能计算出施入氮(肥料氮)在土壤中的残留量及吸收利用情况。随着同位素标记技术的发展,N同位素示踪技术能有效克服上述方法的不足, 由于N同位素无放射性, 对人体没有危害, 是一种稳定的示踪剂, 能精准地计算出标记氮的运移去向, 目前该技术已被广泛应用在氮素循环研究。
近年来, 祝海竣等、Pan等、徐明杰等、党廷辉等、石玉等利用N同位素示踪技术分别研究了水稻()、玉米()、小麦()在“土壤-作物”界面的氮素吸收情况; 岳现录等、Domene等、廖先苓等利用N同位素示踪技术分别分析了肉牛、白鼠、山羊在“牧草-家畜”界面的氮素消化情况。可以看出, 前者多侧重于粮食作物对土壤氮的吸收上, 后者多集中在动物对植物性氮的消化上。但是, 牧草不同于粮食作物, 是不能被人直接消化吸收的, 必须经过反刍动物的消化作用, 才能变成被人吸收的动物性蛋白。而牛、羊作为重要的反刍动物, 其体内的动物性蛋白主要来自于牧草, 牧草中植物性氮又主要来源于土壤, 在这个系统中, 氮素作为一个重要的营养元素, 目前少见从“土壤(土)-牧草(草)-家畜(畜)”系统的角度揭示出氮素运移规律的研究。虽然尹浩冰等综述了草地生态系统的氮循环研究, 但主要集中于“土-草”界面的氮素转移规律, 无法明确吸收的植物氮被反刍动物再利用情况; 黄秀声等虽然分析了8种牧草对氮素的吸收和转化效率, 但在“草-畜”界面是以白鼠为研究对象, 不能真正代表反刍动物实际的吸收情况。因此, 研究氮素在“土壤(土)-牧草(草)-家畜(畜)”界面的运移规律, 对提高牧草氮肥利用效率、明确反刍家畜对植物性氮的消化吸收、以及减少施肥造成的环境污染具有重要意义。
高丹草(×)是高粱()与苏丹草()的远缘杂交种, 具有杂交优势强、生物产量高、饲用品质优、抗逆性强、多茬利用特性, 在农区逐渐受到广大种养户的青睐。近年来, 关于高丹草施氮肥研究主要集中在施氮肥对高丹草产量、品质及环境的影响方面, 尚少见从“土-草-畜”系统研究高丹草氮素运移规律的报道。基于此, 本研究采用N同位素标记技术, 通过施入N标记尿素, 从“土壤残留-植物吸收-动物消化”等3个层面, 探讨了施入氮在高丹草“土-草-畜”界面的运移规律, 研究结果可为高丹草合理施肥、草食家畜对氮素的消化吸收提供理论依据, 为高丹草绿色生产及利用提供技术支撑。
1 材料与方法
1.1 试验地自然概况
试验地位于河北省衡水市的河北省农林科学院旱作农业研究所试验站(E115°42′, N37°44′), 海拔高度20 m, 多年平均气温12.6 ℃, 年平均降水510 mm, 其中70%降水集中在7−8月。无霜期206 d。试验田土壤为黏质壤土, 0~60 cm基础土壤养分: pH 8.25, 全盐量0.93 g∙kg, 有机质13.66 g∙kg, 全氮0.93 g∙kg, 碱解氮58.12 mg∙kg, 速效磷7.61 mg∙kg, 速效钾159.78 mg∙kg。
1.2 试验材料
试验用化肥为N标记的尿素(N丰度为10.17%), 由上海化工研究院生产; 普通尿素(N含量为46.4%), 由沧州大化股份有限公司生产; 磷肥为过磷酸钙肥(PO含量为16%), 由云南个旧大通磷化工有限公司生产。试验所用的高丹草品种为‘冀草2号’, 是由河北省农林科学院旱作农业研究所选育的国审品种, 品种登记号为393, 该杂交种的突出特性: 1)具有光周期敏感特性, 在长日照条件下表现为长时间的营养生长, 短日照条件下营养生长时间变短; 2)保持A型高粱雄性不育的特点, 自交后代不结实。
1.3 试验设计
试验于2015年6−10月在河北省农林科学院旱作农业研究所试验站进行。N标记施肥试验划分5个施肥水平, 纯N量分别为0 kg∙hm、90 kg∙hm、180 kg∙hm、270 kg∙hm和360 kg∙hm, 分别用N0、N90、N180、N270和N360表示, 其中N处理为试验对照(CK)。各N施肥处理的磷肥(PO)施用量为180 kg∙hm, 作为底肥一次性施入。采用随机区组设计, 3次重复, 共计15个小区, 每小区长4.5 m, 宽2.4 m, 总面积10.8 m, 每区种6行高丹草, 南北方向播种, 行距40 cm, 株距15 cm。
N施肥试验采用“小区套微区”设计进行布局, 具体为: 在每试验小区的中间设置为微区, 微区长1.5 m, 宽0.8 m, 面积1.2 m(图1), 施肥时,N标记尿素施在微区; 同时, 在微区四周保留与微区等长、等宽的区域作为保护区(图1), 保护区施肥用普通尿素来替代N标记尿素; 微区和保护区之间没有隔离, 而是利用等渗原理, 同时对微区和保护区进行施肥, 可保证N标记肥料均匀施在微区。同时为防止不同小区之间的肥料渗透, 在各小区之间均留有间隔1.5 m的观察道。
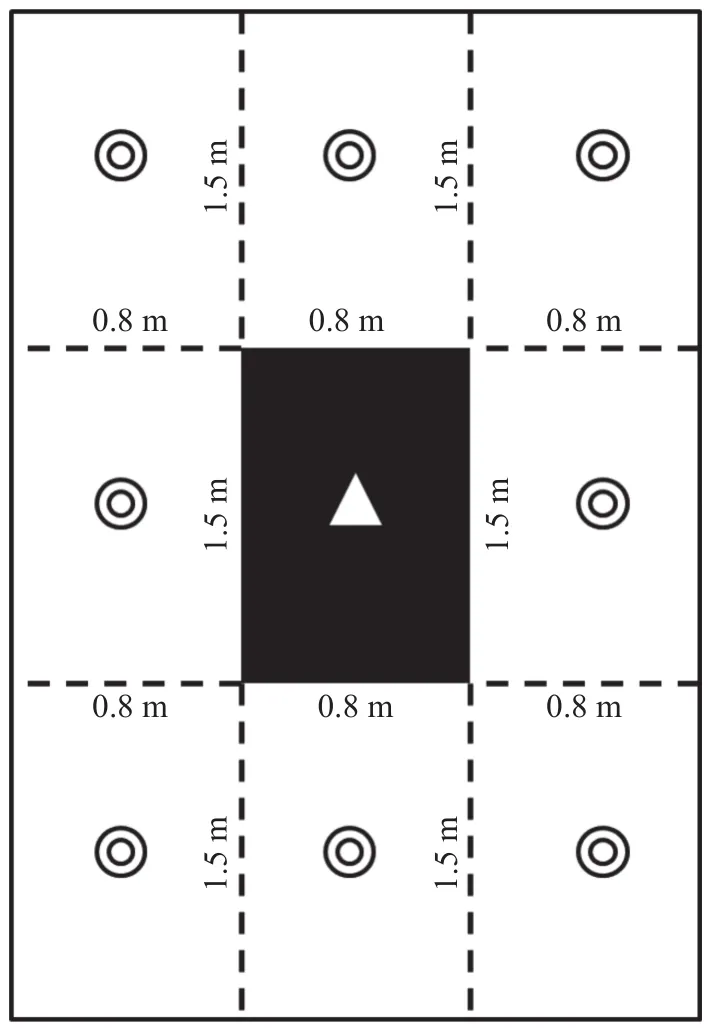
图1 田间小区平面示意图(黑色部分为微区, Δ表示施15N尿素; 白色部分为保护区, ◎表示施普通尿素)Fig.1 Schematic diagram of field plot (black part was micro area, “Δ”means application of 15N urea; white part was protected area, “◎”means application of common urea)
N标记尿素分2次施入, 每次各施50%。第1次作为底肥在播种当天施入, 施肥时在去除试验小区地上杂物、保证平整的基础上, 将N标记尿素均匀撒施微区的土壤表面, 然后翻耕, 与耕作层(0~20 cm土壤)混合均匀; 第2次作为追肥在拔节期(播种后44 d, 株高70 cm左右)施入, 追肥时将N标记尿素溶解于水中, 形成水溶液, 灌入微区, 同时在微区四周的保护区灌溉普通尿素的水溶液, 灌水量按300 m∙hm计算。
试验采用造墒播种, 灌水量为750 m∙hm, 于2015年6月11日点播种植, 播后苗前采用38%莠去津悬浮剂均匀喷施地表防除杂草。
分别于8月19日、10月27日进行第1、2茬草的刈割, 第1茬草刈割时为拔节后期, 第2茬草为孕穗期, 刈割时留茬高度15 cm。每次刈割时, 测定微区株高、茎叶比、草产量等主要农艺性状; 同时, 在每个微区取健壮、长势均匀的单株5株, 茎、叶分开, 自然晾晒成风干样品, 用于测定植物样品中全N和N丰度。在第2茬草刈割后, 对所有处理小区的微区0~1 m的土层取样, 按每0.2 m分层取样, 阴干后将每层土样等量混合, 用于测定土壤样品中全氮含量和N丰度。
为进一步探讨氮在草食动物(肉羊、奶牛)中的运移规律, 在第1茬草刈割后, 收集每个处理3个重复的微区的高丹草鲜样, 用体积为25 L的容器进行青贮发酵, 青贮密度为700 kg∙m, 然后用高丹草青贮草进行肉羊、奶牛的消化率试验, 采用半体内法(瘤胃尼龙袋法)进行。肉羊消化试验选用3只体况正常、健康的装有永久瘘管的绵羊, 将样品放置在瘘管羊瘤胃内进行降解, 48 h取出, 收集残留样品, 测定残留样品的全氮含量和N丰度。奶牛消化试验选用3头装有永久瘘管的处于泌乳中后期的荷斯坦奶牛, 体重650 kg左右, 48 h取出样品, 收集消化后剩余的高丹草残留样品, 测定残留样品的全氮含量和N丰度。
1.4 测定指标与方法
1.4.1 农艺性状测定
株高: 每次刈割时, 测量从地面到植株新叶最高部位的绝对高度; 茎叶比: 在施N标记肥料的微区中选代表性的植株5株, 人工茎、叶分开, 称其鲜重, 自然风干后再称其干重, 可计算出鲜干比和茎叶比;草产量: 测定时先刈割微区四周保护区的高丹草, 然后对微区全部进行刈割测定, 再通过鲜干比折算成干草产量。
1.4.2 土壤养分测定
试验播种前, 采集每个微区0~60 cm的土壤样品, 测定土壤基础养分, 具体取样时, 每20 cm取一层, 阴干后将每层土样等量混合后测定。采用比色法测定土壤的pH, 采用电导率法测定土壤全盐量, 利用稀释热法测定土壤有机质含量, 利用凯氏定氮法测定土壤全氮含量, 碱解氮含量采用扩散法测定, 速效磷含量利用0.5 mol∙LNaHCO浸提法测定, 利用乙酸铵浸提火焰光度法测定速效钾含量。
采用凯氏定氮法测定植物、土壤以及反刍动物消化后样品的全氮含量; 采用ZHT-03型质谱仪测定植物、土壤以及反刍动物消化后样品的N丰度。并计算氮素利用率、土壤残留率、损失率、分配率等相关指标。
1.5 氮素转移计算方法
1.5.1 “土-草”界面氮素运移计算方法


1.5.2 “草-畜”界面氮素运移计算方法

1.6 数据处理
试验旨在明确施入氮在“土-草-畜”界面的运移规律, 通过以下方式实现: 首先计算不同N施肥处理下“土-草”界面N的植物利用率、土壤残留率、损失率, 以及“草-畜”界面N的动物消化率、利用率、残留率值; 然后再将同一界面不同N施肥处理下的上述指标值进行平均, 得出施入氮在不同界面下的运移分配比例; 在此基础上, 明确施入氮在“土-草-畜”界面的运移规律。其中“土-草”界面施入氮的植物利用率、土壤残留率、损失率, 以及施入氮在高丹草茎、叶部位的分配率的计算依据1.5.1部分; “草-畜”界面氮素运移率值依据1.5.2部分计算, 通过计算反刍动物对N高丹草动物消化率, 再进一步换算出反刍动物对施入氮的利用率和残留率。
运用Excel 2007软件计算不同N施肥处理下的高丹草农艺性状值以及施入氮的植物利用率、土壤残留率、损失率、动物利用率等数据的平均值和标准差, 采用SPSS 18.0软件“General Linear Model→Multivariate”进行不同N施肥处理的方差分析; 表中所列数据均用“平均值±标准差”表示。
2 结果与分析
2.1 氮肥对高丹草主要农艺性状的影响
当第1茬草生长69 d后(8月19日)刈割, 不同施肥处理下株高、茎叶比无显著差异; 干草产量在N180处理下为最高, 显著高于CK和N90处理(<0.05), 但与N270、N360处理间的干草产量无显著差异(表1)。第2茬草生长67 d后(10月27日)刈割, 不同施肥处理下株高、茎叶比以及干草产量无显著差异。分析全年干草产量得出, 在N180处理下的总干草产量最高, 显著高于CK和N90处理(<0.05), 但与N270、N360处理间的干草产量无显著差异。

表1 不同15N施肥处理对高丹草不同茬次主要农艺性状的影响Table1 Effect of different 15N treatments on main agronomic characters of Sorghum bicolor×S. sudanense
2.2 氮肥在高丹草“土-草”界面的运移规律
2.2.1 不同施氮处理下施入氮的植物利用率、土壤残留率以及损失率
随着N施肥量的增加, 高丹草植株对施入氮的利用率呈“先增加后降低”趋势, 其中在N180处理下N利用率最高, 为28.44%, 显著高于N360处理(<0.05), 但与N90、N270处理间无显著差异(图2)。随着N施肥量的增加, 0~1 m土壤N残留率呈先增加后降低趋势, 在N180处理下,N元素的残留率达最高, 为28.24%, 显著高于N360处理(<0.05)。随着N施肥量的增加, 施入氮的损失率呈先降低后增加趋势, 在N180施肥处理下,N元素的损失率相对较低, 为43.32%, 但在N360高施肥量处理下,N元素的损失率达60.82%, 与其他处理呈显著差异(<0.05)。

图2 不同15N施用量下高丹草“土-草”界面施入氮的植物利用率、土壤残留率和损失率Fig.2 Plant utilization rate, soil residual rate and loss rate of 15N at the interface of soil-forage of Sorghum bicolor×S. sudanense under different 15N application rates
2.2.2 施入氮在高丹草“土-草”界面的运移分配
平均分析不同N施肥处理下施入氮的利用率、土壤残留率及损失率得出, 施入氮在高丹草“土-草”界面的运移规律为: 施入氮在土壤残留为25.22%, 损失为50.83%, 高丹草吸收23.95%; 总体去向表现为:损失率>土壤残留率>植物吸收率(图2)。
2.2.3 高丹草吸收的氮素在茎、叶的分配
高丹草吸收的施入氮在第1、2茬及全年均以叶中的分配率高于茎, 尤以第2茬最为明显(图3)。而随着N标记肥料的增加, 第1茬、第2茬和全年的茎、叶N分配率在不同施肥处理下均无显著差异。由此表明, 高丹草吸收的氮素在茎、叶的分配率与施肥量多少无关。试验表明, 高丹草营养生长阶段, 叶中的N分配率高于茎。其中叶中施入氮分配率为54.35%, 高于茎中的45.65%。

图3 不同15N施用量下15N在高丹草茎、叶的分配率Fig.3 Distribution rates of 15N in the stem and leaf of Sorghum bicolor×S. sudanense under different 15N application rates
2.3 氮肥在高丹草“草-畜”界面的运移规律
2.3.1 施入氮在肉羊“草-畜”界面的运移规律
不同N处理下, 肉羊对N标记高丹草的消化率及残留率均无显著差异(图4); N180处理下肉羊对施入氮的利用率显著高于N360处理(<0.05), 但与其他处理无显著差异。由图2已知, “土-草”界面中, 施入氮的23.95%可被高丹草吸收利用, 此部分标记的高丹草再经肉羊消化后, 平均分析得出, 施入氮在肉羊“草-畜”界面的运移规律为: 肉羊吸收19.46%, 高丹草残留为4.49% (图4)。综合得出, 施入氮在“土-草-畜(肉羊)”界面的运移规律表现为: 施入氮在土壤残留为25.22%, 在高丹草残留为4.49%, 肉羊吸收19.46%, 损失为50.83%。
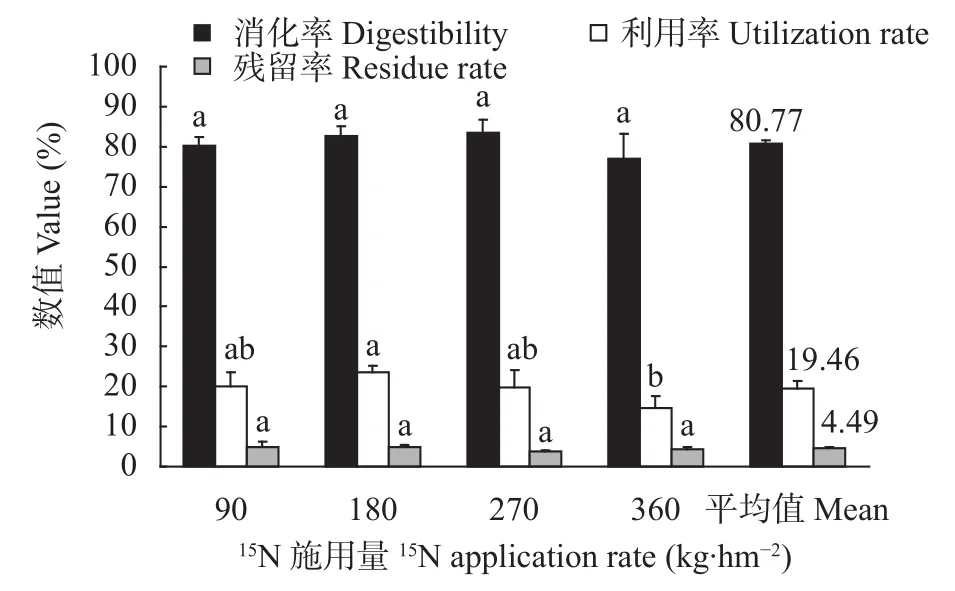
图4 不同15N施用量下“草-畜”界面15N的肉羊消化率、利用率和消化残留率Fig.4 Sheep digestibility, utilization rate and residue rate of15N at the interface of forage-livestock under different 15N application rates
2.3.2 施入氮在奶牛“草-畜”界面的运移规律
N270处理下, 奶牛对N标记高丹草的消化率显著低于N360处理(<0.05), 与其他处理无显著差异;N标记高丹草消化残留率在N180和N270处理下显著高于N360处理, 但与N90处理无显著差异(图5);N180处理下奶牛对施入氮的利用率最高, 但与其他处理间无显著差异。由图2已知, “土-草”界面中, 施入氮的23.95%可被高丹草吸收利用, 此部分标记的高丹草再经奶牛消化后, 平均分析得出, 施入氮在奶牛“草-畜”界面的运移规律为: 奶牛吸收18.17%, 高丹草残留为5.78% (图5)。综合得出, 施入氮在“土-草-畜(奶牛)”界面的运移规律表现为: 施入氮在土壤残留为25.22%, 在高丹草残留为5.78%, 奶牛吸收18.17%, 损失为50.83%。

图5 不同15N施用量下“草-畜”界面15N的奶牛消化率、利用率和消化残留率Fig.5 Dairy cow digestibility, utilization rate and residue rate of 15N at the interface of forage-livestock under different 15N application rates
3 讨论
文献表明, 前人在氮素运移规律研究上多集中在“土壤-作物”或者“牧草-家畜”等单一界面, 主要侧重于氮素利用率的研究; 而牧草是反刍家畜重要的日粮组成, 牧草必须经过反刍动物消化吸收, 才能变成被人吸收的动物性蛋白; 因此研究牧草“土-草-畜”界面的氮素运移规律, 对提高牧草氮肥利用效率、明确反刍动物对植物性氮的消化吸收以及减少施肥造成的环境污染具有重要意义。但相关研究还少有从氮循环角度揭示高丹草“土-草-畜”界面的氮素运移规律。本研究采用N同位素标记法, 通过施入N标记尿素, 可得出“土-草”界面氮素运移比例, 然后再以N标记高丹草为材料, 利用反刍动物模拟消化得出施入N在“草-畜”界面的运移比例; 在此基础上, 揭示了施入氮在高丹草“土-草”界面的运移规律为: 土壤残留25.22%, 损失50.83%, 高丹草吸收23.95%;高丹草再经反刍动物消化后, 施入氮在“土-草-畜(肉羊)”界面的运移规律为: 土壤残留25.22%, 高丹草残留4.49%, 肉羊吸收19.46%, 损失50.83%; 在“土-草-畜(奶牛)”界面的运移规律为: 土壤残留25.22%, 高丹草残留5.78%, 奶牛吸收18.17%, 损失50.83%。
本研究得出高丹草“土-草”N元素的运移规律表现为: 损失率(50.83%)>土壤残留率(25.22%)>植物吸收率(23.95%), 植物吸收率偏低、损失率偏高, 与前人研究结果有一定差异, 可能与该试验的土壤肥力有关, 土壤基础数据显示, 该试验土壤肥力属于中等偏上的肥力, 土壤中氮源丰富, 可能造成高丹草优先吸收土壤氮, 而后再吸收施入的N, 导致高丹草对施入氮的利用率偏低, 而且随着N施肥量的增加, 高丹草对施入氮的利用率表现显著下降趋势, 表明过量施肥不仅没有提高氮肥利用效率, 反而会造成肥料的浪费。土壤残留率与前人研究结果相比较低, 可能是因为前人都是针对一季作物开展的氮素吸收及残留研究, 而本研究中是在收获了两茬高丹草后得到的试验结果, 第2茬草吸收的氮相当于是在第1茬刈割后的基础上, 对第1茬土壤残留N的再吸收, 由此造成土壤中N残留率偏低。然而, 不足之处是未对土壤残留氮的去向进行再研究, 是继续被下茬作物再吸收, 还是经土壤淋溶损失, 还需要进一步研究明确。本试验得出施入氮的损失率偏高, 可能主要原因: 一是与施肥时间和测产周期有关, 本研究采用的是“底施+追施”的方式进行施肥, 追肥是在出苗后35 d进行, 但第1、2茬草的测产时间分别是在出苗后的69 d和136 d完成, 尤其是第2茬草测产时间离追肥期相对较长, 在此过程中, 氮肥挥发或者淋溶损失较多; 二是与微区施肥方式有关, 在微区N施肥上, 将微区和保护区同时施肥, 参考等渗原理, 微区和保护区之间应该没有相互渗透, 因此二者之间并没有采用隔离板阻挡, 而实际在操作中可能会造成N肥料向保护区的渗透, 造成损失; 三是与土壤残留取样有关, 本研究在测定土壤残留时, 取样土层为1 m范围, 也可能由于下渗原因, 1 m以下的土层内也存在N残留, 而这部分残留被算作损失之内。
在“草-畜”界面N元素运移规律研究上, 廖先苓等得出山羊对N标记的稻草消化率为31.16%;Domene等得出Wistar大鼠对菜豆()、豇豆()和豌豆()N的消化率分别为77.6%、84.4%和86.3%; 黄秀声等得出小白鼠对苏丹草N消化率达65.73%;岳现录等得出肉牛对青贮玉米N的吸收和损失率合计为64.0%; Powell等得出奶牛对N标记的饲料消化后, 粪便N残留率为22%~28%, 而对普通饲料添加N 化肥消化后, 粪便中N残留率为64%~78%。可以看出, 前人研究的重点是不同动物对N标记牧草的消化率和残留率的研究, 但均未从“土-草-畜”系统揭示施入氮被反刍动物吸收的情况。基于此, 本研究以N标记高丹草为试验材料, 通过饲喂肉羊、奶牛等反刍动物, 从“土-草-畜”系统解释了施入氮被反刍动物吸收的情况, 结果得出, 施入氮经“土-草-畜(肉羊)”界面转化后, 肉羊对施入氮的利用率为19.46%, 经“土-草-畜(奶牛)”界面转化后, 奶牛对施入氮的利用率为18.17%。不足之处是由于N肥料价格昂贵, 形成的N标记高丹草样品数量少, 难以完成系统的饲喂试验, 因此在计算反刍动物消化率时只能采用瘤胃尼龙袋法模拟进行, 实际上动物瘤胃消化部分并不能完全被家畜吸收, 也可能随粪便代谢一部分, 同时在试验中忽略了高丹草青贮过程中的氮损失量, 由于这两部分的损失未计算, 实际测定的肉羊、奶牛N元素利用率值相对偏高, 在以后试验中还需注意改进。同时, 由于试验条件有限, 本研究重点分析了施入氮在“土-草-畜”界面的运移规律, 未考虑反刍动物代谢后的粪尿还田后被高丹草再吸收过程中的氮素运移规律, 未来还需加强这一过程研究, 为研究种养结合模式中的氮素循环研究提供理论依据。
在不同作物器官氮素分配比例研究上, 晏娟等在水稻、徐明杰等在玉米上的研究得出, 开花期前N主要分配在叶片中, 而花后逐渐向籽粒中转移。晏娟等得出, 水稻开花期之前N主要分配在叶片中, 其分配比例在58%以上; 而花后籽粒中N分配比例在55%以上。徐明杰等得出, 玉米开花期叶片N分配率在46%以上, 花后籽粒中N分配比例在60%以上。马兴华等研究得出, 成熟期氮素在小麦植株中的分配比例为籽粒>茎秆+叶鞘>叶片>穗轴+颖壳, 其中籽粒中的分配比例约80%, 营养器官中的分配比例约20%。上述研究初步得出,N元素转移规律基本表现为开花期主要积累在叶片中, 而随着籽实的形成,N元素逐渐向籽实转移; 而高丹草与粮食作物不同, 它是以收获营养体为主的, 并不需要完成整个生育期; 因此在高丹草营养器官在氮素分配比例与粮食作物是有区别的, 本研究明确了高丹草茎叶器官中的氮素的分配比例。平均分析得出, 高丹草营养生长阶段, 叶中N元素分配率为54.35%, 高于茎中的45.65%。
在海河平原区高丹草合理施氮量研究中, 李源等曾得出该区高丹草合理的施氮量为180~270 kg∙hm, 但在中高等肥力土壤下高丹草是否可减施或隔年(季节)施肥还需进一步研究。本研究结果得出, 随着施肥量增加, 高丹草氮素利用效率并没有显著增加, 最高利用率不足30%, 施肥过多则造成了资源的浪费。由此表明, 在中高等肥力土壤下, 高丹草可减施或隔年(季节)施肥。
4 结论
本试验采用N同位素标记法, 揭示了高丹草“土-草-畜”界面的氮素运移规律。在高丹草“土-草”界面施入氮的运移规律为: 土壤残留25.22%, 损失50.83%, 高丹草吸收23.95%; 施入氮在高丹草茎、叶部位的分配规律为: 叶(54.35%)>茎(45.65%); 高丹草再经反刍动物消化后, 施入氮在“土-草-畜(肉羊)”界面的运移规律为: 土壤残留25.22%, 高丹草残留4.49%, 肉羊吸收19.46%, 损失50.83%; 在“土-草-畜(奶牛)”界面的运移规律为: 土壤残留25.22%, 高丹草残留5.78%, 奶牛吸收18.17%, 损失50.83%。研究结果可为高丹草合理施肥及草畜转化研究提供理论依据。
