外源ATP对摄磷烟草悬浮细胞生长和活性氧代谢的影响
2022-08-05冯汉青王娟娟达晓伟杨德丽庞海龙
冯汉青,王娟娟,达晓伟,杨德丽,庞海龙
(西北师范大学 生命科学学院,甘肃 兰州 730070)
磷作为植物所需的常量元素,是维持植物生长发育的重要物质基础[1].自然界中植物所需的磷通常以磷酸盐形式存在.由于磷是核酸、磷脂类等物质的构成元素,因此无论是自然界的植物还是体外培养的植物组织或细胞在生长过程中均需要较大的磷酸盐供给以维持基本的生长和发育[2].
磷源供应水平的变化能够引起细胞的多种生理学反应[3].研究发现,细胞磷供应量决定着核酸、蛋白质、花色苷、苯丙素、花青素等初级和次级代谢物的合成水平[4-5].另有研究揭示,环境中可摄取磷酸盐量减少时会明显影响细胞呼吸途径[6]、糖酵解途径的运行,并影响到植酸酶、抗氧化酶、酸性磷酸酶等植物代谢相关酶的活性[7-8].
三磷酸腺苷(ATP)主要在细胞内产生,是生物体内的“能量货币”.而最近的研究发现,动物、植物以及微生物细胞均可以将细胞内ATP通过多种方式释放到细胞外基质中,使得细胞外也存在着一定水平的ATP,即胞外ATP(eATP)[9].对动物细胞研究发现,胞外ATP可以结合跨膜受体蛋白,作为信号分子调节细胞生长及免疫等生理过程[10-11].胞外ATP也是植物中重要的信号分子[12],调控了植物细胞活性氧(ROS)及钙离子的水平、生长发育、防御相关基因表达以及植物对于逆境的响应等[13-14].另有研究报道,胞外ATP过多的积累以及胞外ATP的去除均能诱导细胞死亡,说明胞外ATP的动态平衡在植物细胞活性的调控中也扮演着重要的角色[15].因此,基于胞外ATP的上述功能,外源ATP可以作为调控剂影响植物的多种生理学反应.
如上所述,磷酸盐的供应会影响到细胞的生长和生理学状态[16-17].但目前为止,在缺磷及正常磷供应下外源ATP对于细胞生长和活性氧代谢的影响以及差异尚不明确.故文中以烟草悬浮细胞为材料,探讨了在缺磷及正常磷供应下胞外ATP对细胞生长和抗氧化性能的影响,以期进一步为胞外ATP调控植物生长和抗氧化代谢的机理研究提供一定的理论依据.
1 材料与方法
1.1 实验材料
所用烟草悬浮细胞(NicotianatabacumL.cv.Bright Yellow-2)由香港中文大学姜里文教授馈赠.将烟草悬浮细胞在Murashige-Skoog(MS)液体培养基(添加3%(W/V)蔗糖和0.2 mg·L-1的2,4-二氯苯氧乙酸(2,4-D))中接种,于27 ℃,130 r·min-1黑暗条件下振荡培养.每隔一个生长周期(7 d)吸取10 mL细胞培养物,加入到200 mL新鲜的MS液体培养基中进行传代.吸取生长对数期(第4 d)的烟草悬浮细胞为实验材料进行处理,所有步骤均在无菌条件下进行.
1.2 烟草悬浮细胞的处理
取处于生长对数期(第4 d)的烟草悬浮细胞.用金属网筛过滤后,称取2 g细胞(鲜重)用去离子水洗涤2次后分别进行如下处理:将上述烟草悬浮细胞分别加入到缺磷MS培养(不含KH2PO4)、标准MS培养基(含1.25 mmol·L-1KH2PO4)、添加外源ATP(30 μmol·L-1)的缺磷MS培养基和添加外源ATP(30 μmol·L-1)的标准MS培养基.所有处理浓度均为细胞悬浮液中的终浓度,终体积为20 mL.将上述不同培养基下生长的细胞在27 ℃黑暗条件下振荡培养72 h.
1.3 H2O2的测定
H2O2含量采用硫酸钛法测定[18].称取0.2 g细胞,加入1 mL 4 ℃预冷丙酮,冰浴研磨成匀浆,于4 ℃,10 000 r·min-1离心10 min;取0.7 mL上清液并加入0.1 mL 5%硫酸钛和0.2 mL浓氨水,在4 ℃,10 000 r·min-1下离心10 min.弃上清后用1 mL 2 mol·L-1浓硫酸溶解沉淀,在OD415处测定吸光度.根据标准曲线计算H2O2含量.
1.4 细胞活力测定
细胞活力采用伊文思蓝法测定[19].称取0.2 g细胞,加入1 mL磷酸缓冲液摇匀后,再加入质量分数为0.025% 的伊文思蓝染液0.1 mL,染色8 min.3 000 r·min-1下离心3 min 弃上清,再次加入1 mL 的磷酸缓冲液,于3 000 r·min-1下离心3 min弃上清. 多次重复此步骤以洗去未结合细胞的伊文思蓝.对清洗后的细胞加入1 mL质量分数为1% 的十二烷基磺酸钠溶液(溶剂为体积分数50%的甲醇),混匀后于50 ℃下水浴30 min,使细胞裂解,并于10000 r·min-1下离心10 min,取0.5 mL上清液,用蒸馏水稀释至3 mL,在OD595处检测吸光值.
1.5 细胞鲜重测定
细胞培养72 h后,充分摇起锥形瓶中的细胞,用金属网筛滤去培养基,利用电子天平称量细胞,得细胞鲜重.
1.6 超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)的测定
1)酶液提取.称取0.2 g烟草悬浮细胞于预冷研钵中,加入预冷磷酸缓冲液(0.05 mol·L-1pH=7.8)及少许石英砂,充分研磨后加入缓冲液至终体积为1 mL.于4 ℃,10 000 r·min-1离心10 min.取上清即为待测粗酶液.
2)POD活性用愈创木酚法测定[20].POD酶活性以OD470每分钟变化0.01为一个酶活单位(U).SOD活性测定用NBT光化学还原法[21],SOD酶活性以抑制NBT光化学还原的50%为一个酶活单位(U).CAT活性测定用紫外吸收法[22],CAT 酶活性以OD240每分钟变化0.1为一个酶活单位(U).
1.7 统计学分析
所得结果为至少3次实验的平均值,实验结果均用平均值±标准差表示,数据采用双总体t检验,在P<0.05水平上具有差异显著性.
2 结果与分析
2.1 缺磷及正常磷供应下外源ATP对烟草悬浮细胞活性和生长的影响
结果显示,缺磷下生长的烟草细胞活性是正常磷供应下细胞活性的1.2倍.对于缺磷及正常磷供应下生长的细胞分别加入外源ATP后发现,外源ATP使得缺磷下细胞的活性降低了13%,但外源ATP使得正常磷供应下生长的细胞活性升高了10%(效果未达到显著性水平, 图1(a)).
测定细胞的鲜重发现,正常磷供应下生长的细胞的鲜重是缺磷下的1.2倍.对于缺磷及正常磷供应下生长的细胞分别加入外源ATP后发现,外源ATP使缺磷下的细胞鲜重增加了1.5倍,而正常磷供应下生长的细胞鲜重增加了1.3倍(图1(b)).
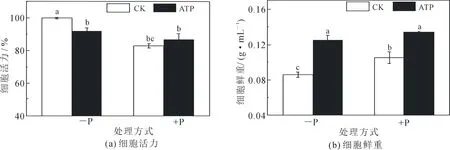
注:-P,缺磷供应;+P,正常磷供应;CK,未进行外源ATP处理的细胞;ATP,进行外源ATP处理的细胞.不同字母表示在P<0.05水平上具有显著性差异,下同.
2.2 缺磷及正常磷供应下外源ATP对烟草悬浮细胞H2O2水平的影响
通过检测细胞H2O2含量发现,正常磷供应下生长的细胞H2O2含量是缺磷下生长细胞H2O2含量的3.8倍.对缺磷及正常磷供应下生长的细胞分别添加外源ATP,结果显示外源ATP使缺磷下生长的细胞中H2O2含量显著性升高了28%,而使正常磷供应下生长的细胞中的H2O2含量显著降低了23%(图2).
2.3 缺磷及正常磷供应下外源ATP对烟草悬浮细胞POD、SOD、CAT活性的影响
缺磷下生长的细胞的POD活性是正常磷供应下生长细胞POD活性的7.5倍.分别对缺磷及正常磷供应下生长的细胞添加外源ATP,发现外源ATP使得缺磷下生长的细胞的POD活性显著性降低了60.2%,而使正常磷供应下细胞POD活性显著性上升了7.4倍(图3(a)).
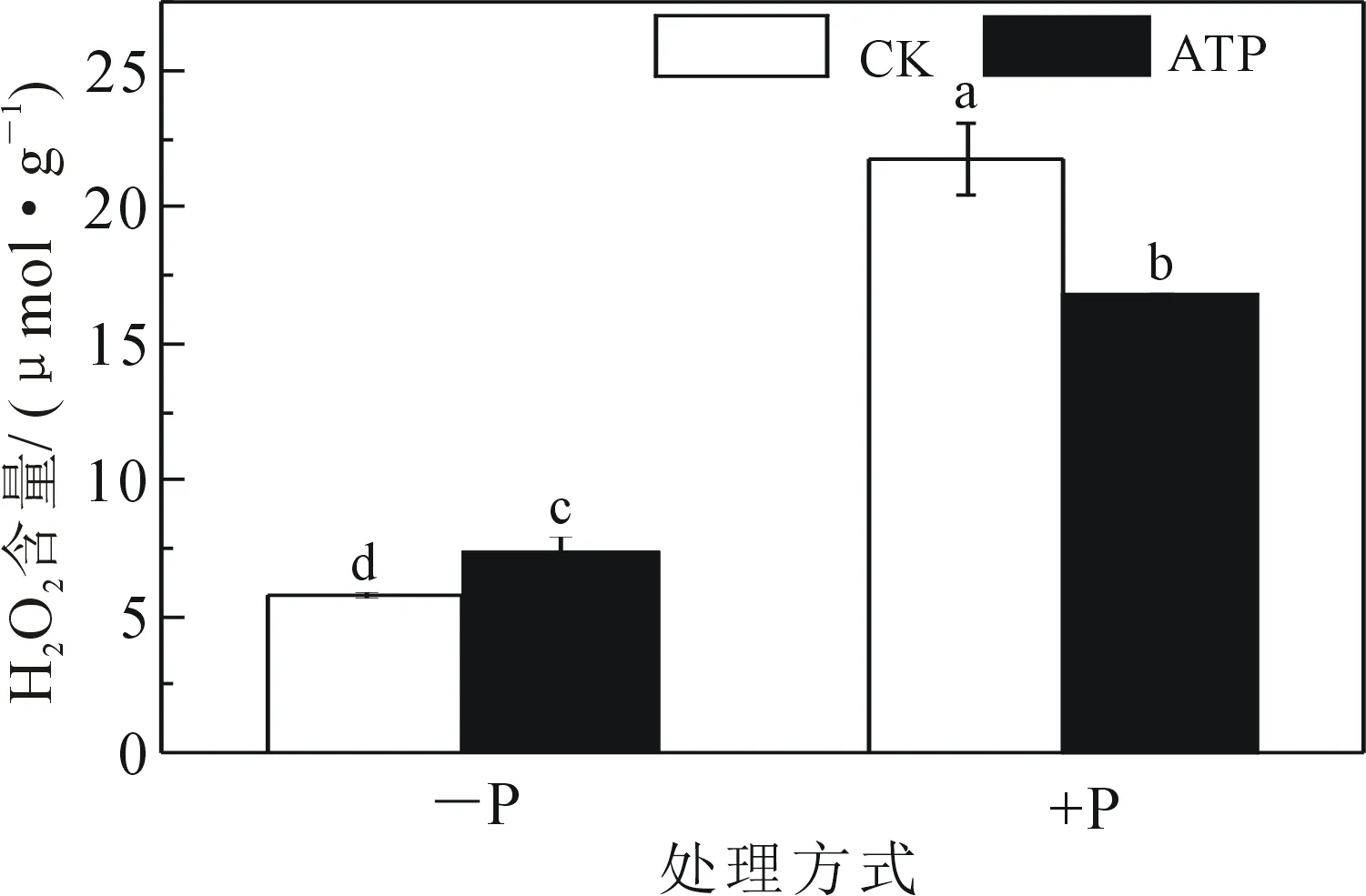
图2 不同处理下BY-2烟草悬浮细胞H2O2含量的变化
SOD活性测定结果显示,缺磷下生长的细胞的SOD活性略高于正常磷供应下生长的细胞,但未达到显著性差异.添加外源ATP使得缺磷及正常磷供应下生长的细胞SOD活性均有所下降(图3(b)),但均未达到显著水平.
CAT活性测定结果显示,缺磷下生长的细胞的CAT活性是正常磷供应下生长细胞的1.6倍.外源ATP使缺磷下生长的细胞的CAT酶活性显著性上升了2.5倍,而使正常磷供应下生长细胞的CAT活性显著性上升了3倍(图3(c)).
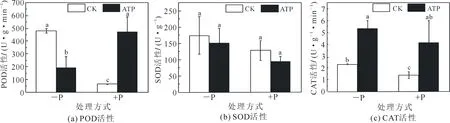
图3 不同处理下POD,SOD,CAT活性的变化
2.4 外源ATP对摄磷烟草悬浮细胞生长和活性氧代谢影响分析
如图4,较之细胞的活力,细胞的鲜重对外源ATP更加敏感;而在活性氧代谢过程中,较之细胞SOD活性,细胞的H2O2含量、POD活性以及CAT活性对外源ATP更加敏感.对于缺磷及正常磷供应下生长的细胞,外源ATP对于细胞鲜重、SOD活性以及CAT活性具有相似的影响;而外源ATP对于细胞活力、H2O2含量、POD活性则具有相反的影响;其中,外源ATP对POD活性的影响最为明显.
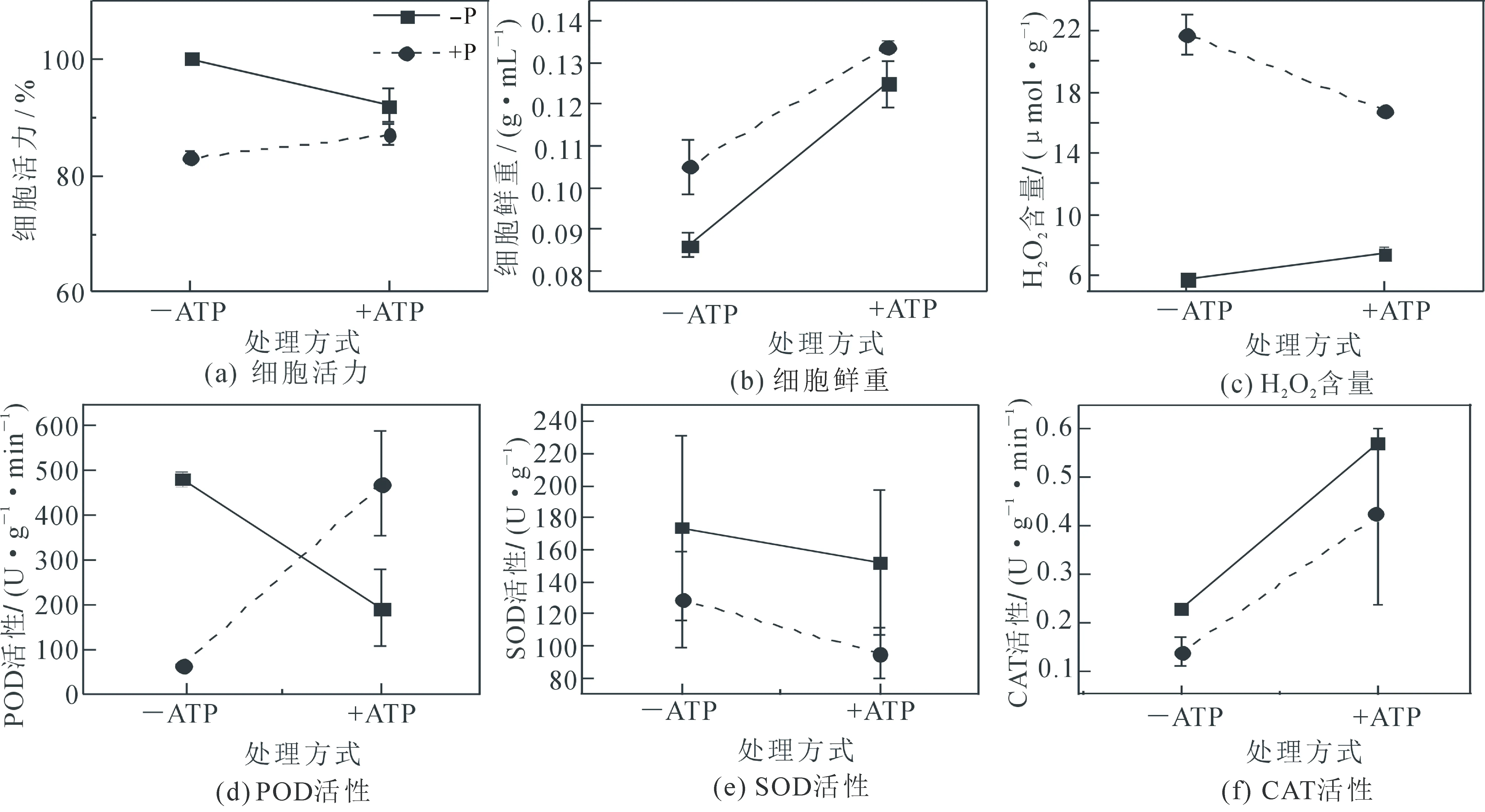
图4 外源ATP对摄磷细胞生长和活性氧代谢的影响趋势
3 讨论
作为一种大量营养元素,环境中磷素的供应影响着植物的生长状态和代谢水平[23].本研究发现,较之缺磷下生长的细胞,正常磷供应下生长的细胞活性降低但鲜重增加.说明在有磷酸盐供应时,伴随着细胞的生长,细胞膜会产生一定的损伤.
研究表明,外源磷的缺乏能够限制细胞的分裂[27-28].因此,正常磷供应下细胞鲜重的增加可能是磷供应刺激细胞分裂的结果.另有研究发现,在磷酸盐浓度较低时,细胞膜表面质子泵不会被激活;而当有磷酸盐摄入时,细胞质发生酸化以刺激质子泵运输磷[24];而细胞质酸化被认为能够增加膜的透性和细胞损伤[25-26].因此,文中认为在正常磷供应下细胞出现的损伤应当为细胞加速吸收和运输磷过程中细胞质发生酸化所致.
对比缺磷及正常磷供应下生长的细胞对外源ATP的反应(图4a-b)发现,外源ATP能够促进两种细胞鲜重的增长,并显著降低了缺磷生长下细胞的活性.尽管植物细胞无法直接吸收外源ATP,但细胞外ATP受体蛋白或三磷酸腺苷双磷酸酶可感受胞外ATP或水解细胞外ATP产生磷酸盐[29].据此推测,外源ATP增加了细胞磷酸盐的供给,由此增加了细胞的分裂和生长.而对于缺磷生长的细胞而言,外源ATP使得细胞膜表面质子泵从非激活状态转化为激活状态,导致细胞质明显的酸化和较为明显的细胞损伤.
进一步研究揭示,较之缺磷下生长的细胞,正常磷供应下生长的细胞H2O2水平显著上升.Hannah等也发现低磷会导致烟草悬浮细胞ROS水平降低[30].另有研究揭示,缺磷会增加植物抗氧化酶的基因表达和活性[32-36].因此,缺磷下细胞活性氧含量的变化应当和抗氧化酶的变化有关.文中通过测定抗氧化酶发现,正常磷供应下生长的细胞POD、SOD、CAT活性均低于缺磷下生长的细胞,其中POD的活性降低最为显著.据此推测,POD活性的降低是导致正常磷供应下生长细胞H2O2水平上升的主要原因.
早期在动物细胞中发现,细胞外ATP可以诱导ROS产生.继而在植物中也发现,细胞外ATP能够上调ROS的产生[37].有趣的是,外源ATP也可以诱导采后农作物抗氧化酶活性的提高,以缓解ROS积累所导致的采后农作物衰老[39].因此,细胞外ATP对植物细胞ROS代谢的调控尚有一定的争议.
文中也继而研究了外源ATP对缺磷及正常磷供应下生长的细胞ROS代谢的影响.结果显示,对于两种细胞,外源ATP均增加了CAT和SOD的活性;但外源ATP对H2O2水平和POD的活性具有相反的作用(图4(c)~(f)):对于缺磷下生长的细胞,ATP降低了H2O2水平,但增加了POD的活性;对于正常磷供应下生长的细胞,ATP增加了H2O2水平,但降低了POD的活性.这一方面提示,POD活性的变化是决定缺磷及正常磷供应下生长的细胞H2O2水平变化的主要原因;另一方面则表明,外源ATP对于植物细胞H2O2的调节作用和细胞磷酸盐的供给水平有关:在正常磷供应下,外源ATP能够通过下调POD的活性而增加H2O2的产生;而在缺磷状态下,外源ATP能够通过上调POD的活性而降低H2O2的产生.这可能解答了目前对于细胞外ATP对植物细胞活性氧代谢调控作用的争议:对于在磷供应正常下生长的植物,外源ATP能够促进H2O2的产生;而对于采后的农作物,由于已经无法从环境中摄取磷酸盐,外源ATP则表现为抑制H2O2的产生.
综上所述,在缺磷及正常磷供应下外源ATP能调节细胞摄磷过程中细胞的生长和ROS代谢,且其调控作用和细胞的磷源供应有关.
