窝儿七的光合作用与环境因子间的关系研究*
2022-08-01孙涛雷瑞祥刘永裴青青
孙涛 雷瑞祥 刘永 裴青青
(陕西中医药大学,陕西 咸阳 712046)
窝儿七为小檗科山荷叶属植物南方山荷叶(DiphylleiasinensisH.L.Li)的根及根茎,产于湖北、陕西等地,生长于海拔1880~3700米的落叶阔叶林、针叶林、竹丛或灌丛下。根茎具有清热、凉血、活血、止痛以及泻下作用。主治腰腿疼痛、风湿性关节炎、跌打损伤、月经不调等症[1]。窝儿七中主要化学成分有鬼臼毒素、去氢鬼臼素、β-谷甾醇、山柰酚糖苷、山荷叶素糖苷、鼠李柠檬素、鼠李素和闭木花酮、棕榈酸、3-O-甲基-槲皮素等[2]。其中的黄酮类化合物具有直接抑菌、协同抑菌及抑制细菌毒性等作用[3],鬼臼毒素类物质具有良好的抗肿瘤和抗免疫活性[4],因此窝儿七药材具有广阔的应用前景。
如何提高中药材的品质与产量一直是当前的研究热点,目前对窝儿七的化学成分活性,有效成分的含量、药效等方面研究较多[5-6],未见有关其光合作用研究报道,光对植物的形态建成和生殖器官的发育影响很大,光合作用的产物是药用植物生长发育的物质基础,光合作用强弱直接决定药用植物的产量[7],所以研究窝儿七的光合作用有利于了解其生长规律。本文以小檗科植物窝儿七的叶片为研究材料,对窝儿七净光合速率日变化及其环境因子的相互关系进行研究,揭示环境因子对窝儿七叶片的光合特性的日变化规律,为选择合理的栽培环境和栽培种源提供依据。
实验研究在2018年7月25日,在太白县塘口村进行试验,塘口村处于秦岭鳌山北麓,由栓皮栎、蒙古栎、华山松和桦树形成落叶阔叶林,全年降雨在1000 mL左右,温度-30 ℃~20 ℃之间,窝儿七生长于林下阴湿环境中。试验材料选择上层功能叶片,连续晴天之后进行相关光合试验。材料经陕西中医药大学中药资源学教研室王西芳教授鉴定为小檗科植物南方山荷DiphylleiasinensisH.L.Li。
1 方法
1.1光合参数日变化测定 采用便携式光合测定仪(TARGAS-1,USA),从8∶00~18∶00每隔1 h测定自然条件下窝儿七叶片的光合日变化。测定的指标主要有:净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)以及主要环境因子,包括光合有效辐射(PAR)、大气温度(Ta)、空气相对湿度(RH)、大气CO2浓度(Ca)等。测定前选择5株健壮植株,选取生长部位相同、向阳角度一致的5片叶片进行活体测定,测定时保持叶片自然生长角度不变,结果为5次测定的平均值。同时计算窝儿七叶片水分利用率(WUE=Pn/Tr)及气孔限制值(Ls=1-Ci/Ca)。
1.2光响应曲线的测定 被测叶片在自然光强下适应2 h,于晴朗天气的上午9∶00~11∶00测定光响应曲线,温度和CO2浓度采用自然条件。光合有效辐射(PAR)采用便携式光合测定仪内置红蓝光源控制,光照强度梯度设为0、50、100、200、300、400、600、800、1000、1200、1500、2000 μmol·m-2·s-1,叶片选择同上。
1.3数据处理 采用Excel 2007和SPSS 20.0软件进行正交数据处理分析。
2 结果和分析
2.1光合有效辐射(PAR)日变化 由图1可知,窝儿七叶片光合有效辐射(PAR)日变化的规律呈现“单峰”曲线。8∶00~9∶00光合有效辐射呈现快速上升状态,9∶00=~10∶00=光合有效辐射没有变化,10∶00~12∶00光合有效辐射快速上升,在12∶00达到最大值(1631 μmol·m-2·s-1),12∶00~14∶00光合有效辐射缓慢降低,14∶00~17∶00光合有效辐射快速降低,18∶00时测得光合有效辐射最小值(213 μmol·m-2·s-1)。
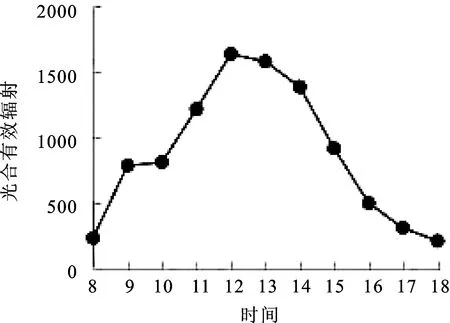
图1 光合有效辐射(PAR)日变化
2.2大气温度(Ta)日变化 温度对植物的生长发育至关重要,过高或过低的温度会对植物的光合特性产生显著影响[8]。由图2可知,大气温度(Ta)随着时间的不断推移,变化趋势与PAR变化基本一致,其变化范围在15.2~31.3℃之间,8∶00~12∶00大气温度快速上升并于12∶00达到最大值,12∶00~15∶00大气温度保持平稳状态,15∶00~18∶00大气温度快速下降。
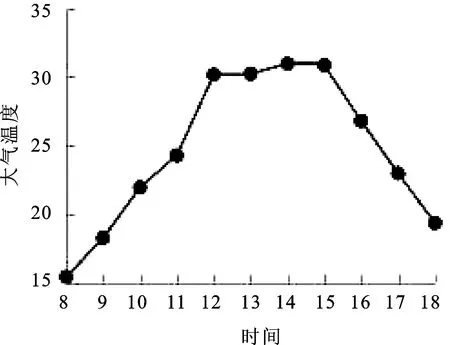
图2 大气温度(Ta)日变化
2.3空气相对湿度(RH)日变化 研究表明,空气相对湿度较高的早晨光合速率不高,认为这除了与光照有关外,还与空气相对湿度大,蒸腾速率低,不利于光合产物在植株体内的运输有关[9]。由图3可知,空气相对湿度(RH)的变化趋势与光合有效辐射和大气温度的变化趋势呈现负相关,8∶00=空气相对湿度最大(58.34%),8∶00=~13∶00=随着大气温度与光有效辐射的不断增大,空气相对湿度不断降低,13∶00=~15∶00=相对平缓,14∶00=测得其最小值(35%),15∶00=~18∶00=随着大气温度与光有效辐射的降低,空气相对湿度快速上升。
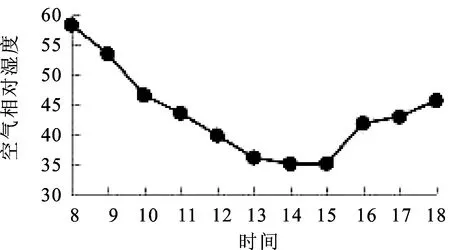
图3 空气相对湿度(RH)日变化
2.4大气CO2浓度(Ca)日变化 如图4分析可以得出,大气CO2浓度(Ca)的变化趋势与空气相对湿度变化趋势类似,其变化范围为385.94~469.52 μmol·m-2·s-1,8∶00=大气CO2浓度最大,8∶00=~11∶00=大气CO2浓度逐渐降低,并于11∶00=达到最小值(385.94 μmol·m-2·s-1),11∶00=~13∶00=大气CO2浓度逐渐上升,变化速率相对缓慢,13∶00=~18∶00=大气CO2浓度变化不大,保持平稳。CO2是植物生长发育和生理活动的关键因子,研究表明,大气CO2浓度升高会增强植物光合作用[10]。
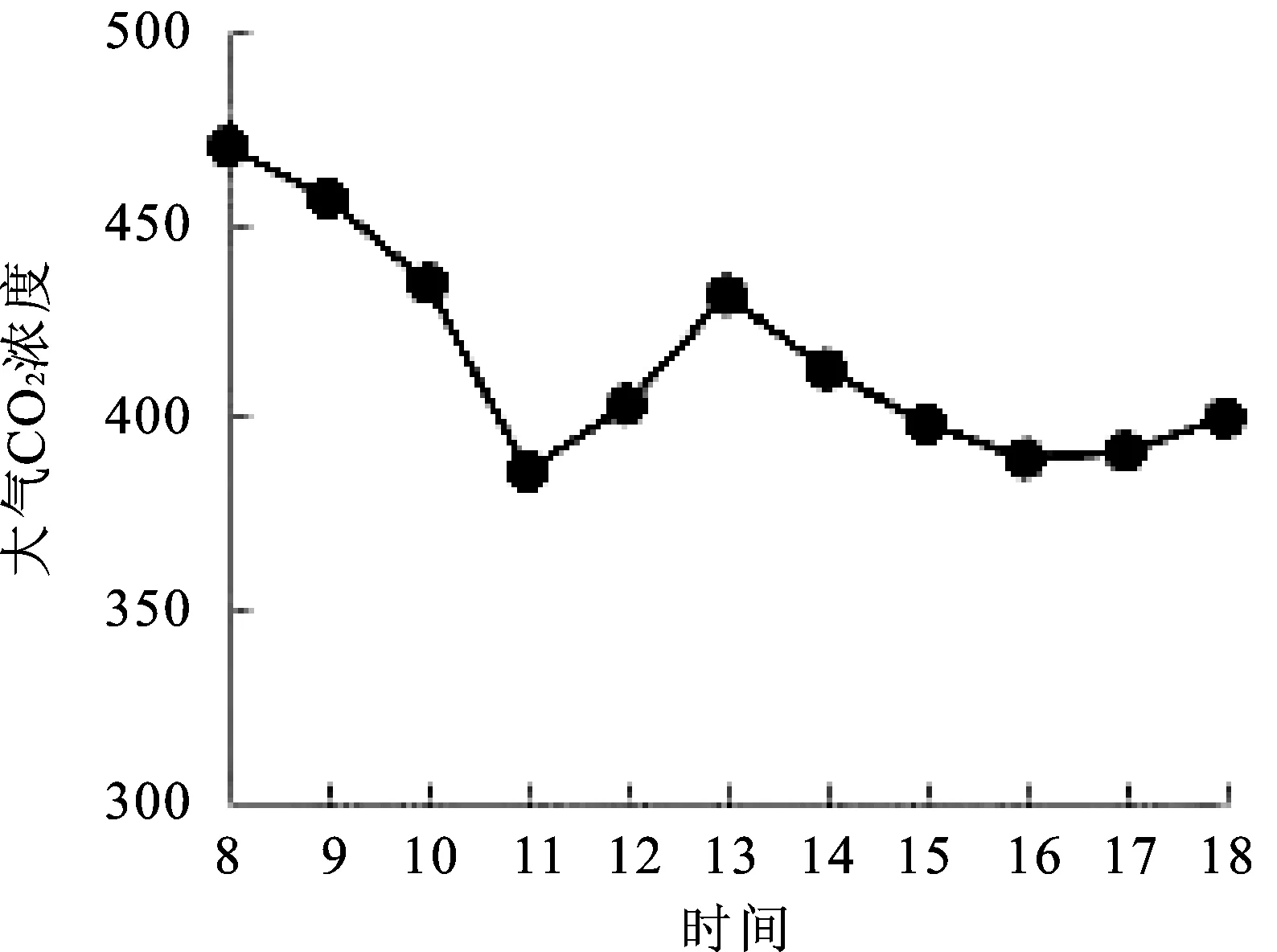
图4 大气CO2浓度(Ca)日变化
2.5净光合速率(Pn)日变化 净光合速率是表示光合作用强弱变化的核心指标之一[11]。由图5可知,窝儿七叶片净光合速率Pn日变化呈“单峰”曲线,8∶00=~10∶00=净光合速率呈上升趋势,8∶00=~13∶00=净光合速率上升减慢,并于13∶00=达到最大值16.21 μmol·m-2·s-1,13∶00=~18∶00=净光合速率快速下降,于18∶00=到达最小值8.22 μmol·m-2·s-1。
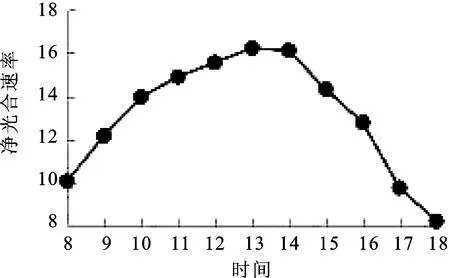
图5 净光合速率(Pn)日变化
2.6气孔导度(Gs)日变化 气孔是植物水分和CO2气体交换的重要通道,气孔导度是决定植物光合作用强度和水分蒸腾强度的重要因素[12]。由图6可知,窝儿七叶片气孔导度与净光合速率日变化规律基本一致,以“单峰”的形式表现出来。8∶00=~9∶00=气孔导度呈现缓慢上升趋势,9∶00=~11∶00=快速上升并达到峰值0.56 μmol·m-2·s-1,从11∶00=~18∶00=一直下降并达到最小值0.12 μmol·m-2·s-1,晚上18∶00=的气孔导度明显小于早8∶00=的气孔导度数值。
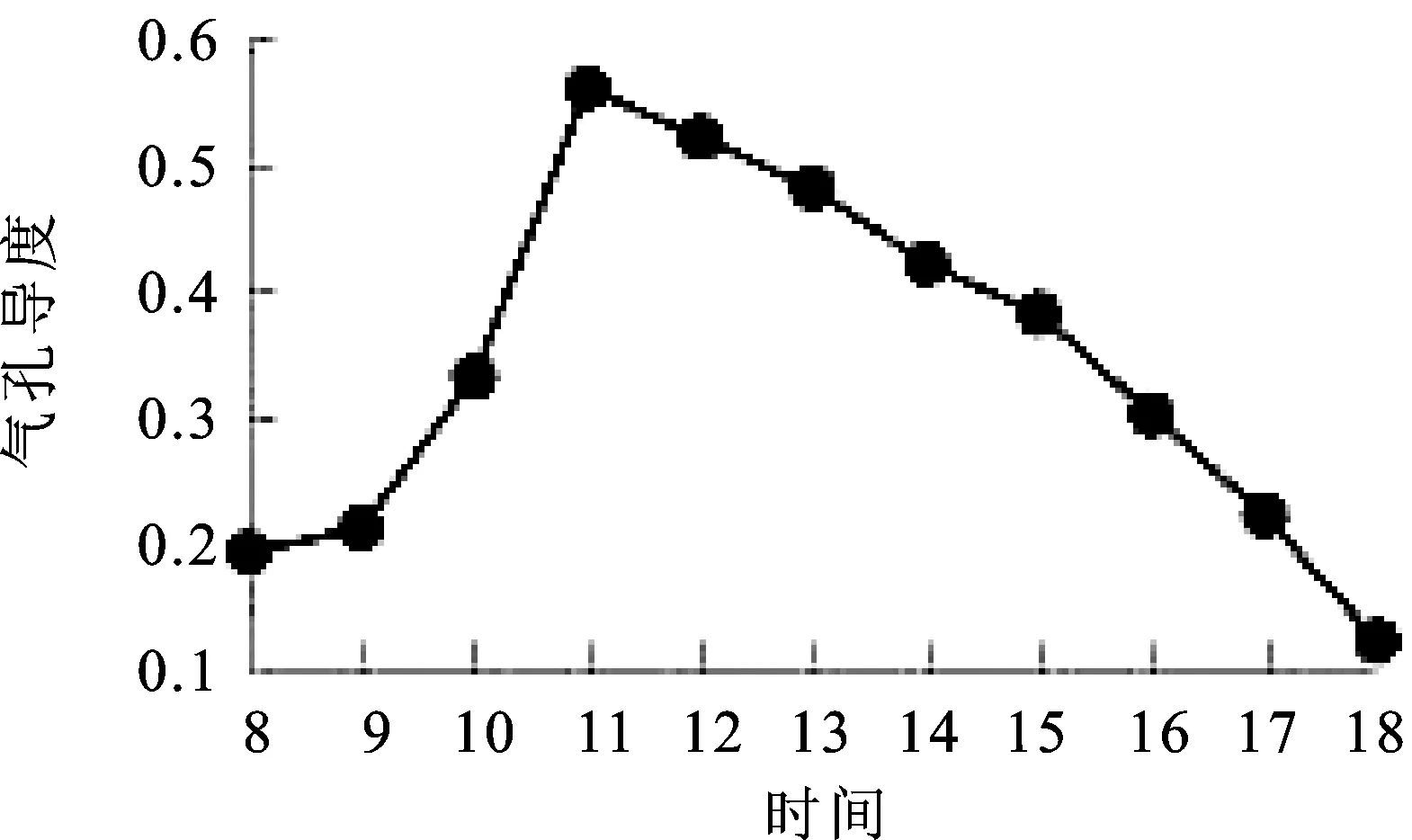
图6 气孔导度(Gs)日变化
2.7蒸腾速率(Tr)日变化 蒸腾作用主要涉及植物水分的吸收和有机物的运输过程,是营养物质在植物体内运输的主要动力,蒸腾速率的大小反映植物吸收和运输能力的强弱[13]。由图7可知,窝儿七叶片蒸腾速率Tr日变化曲线与PAR和Ta的趋势基本一致,也呈“单峰”曲线。8∶00=~13∶00=蒸腾速率持续上升,13∶00=出现高峰值7.30 μmol·m-2·s-1,13∶00=~14∶00=蒸腾速率呈缓慢下降趋势,14∶00=~18∶00=蒸腾速率快速下降,18∶00=到达最小值1.98 μmol·m-2·s-1,18∶00=的蒸腾速率稍大于8∶00=的蒸腾速率。
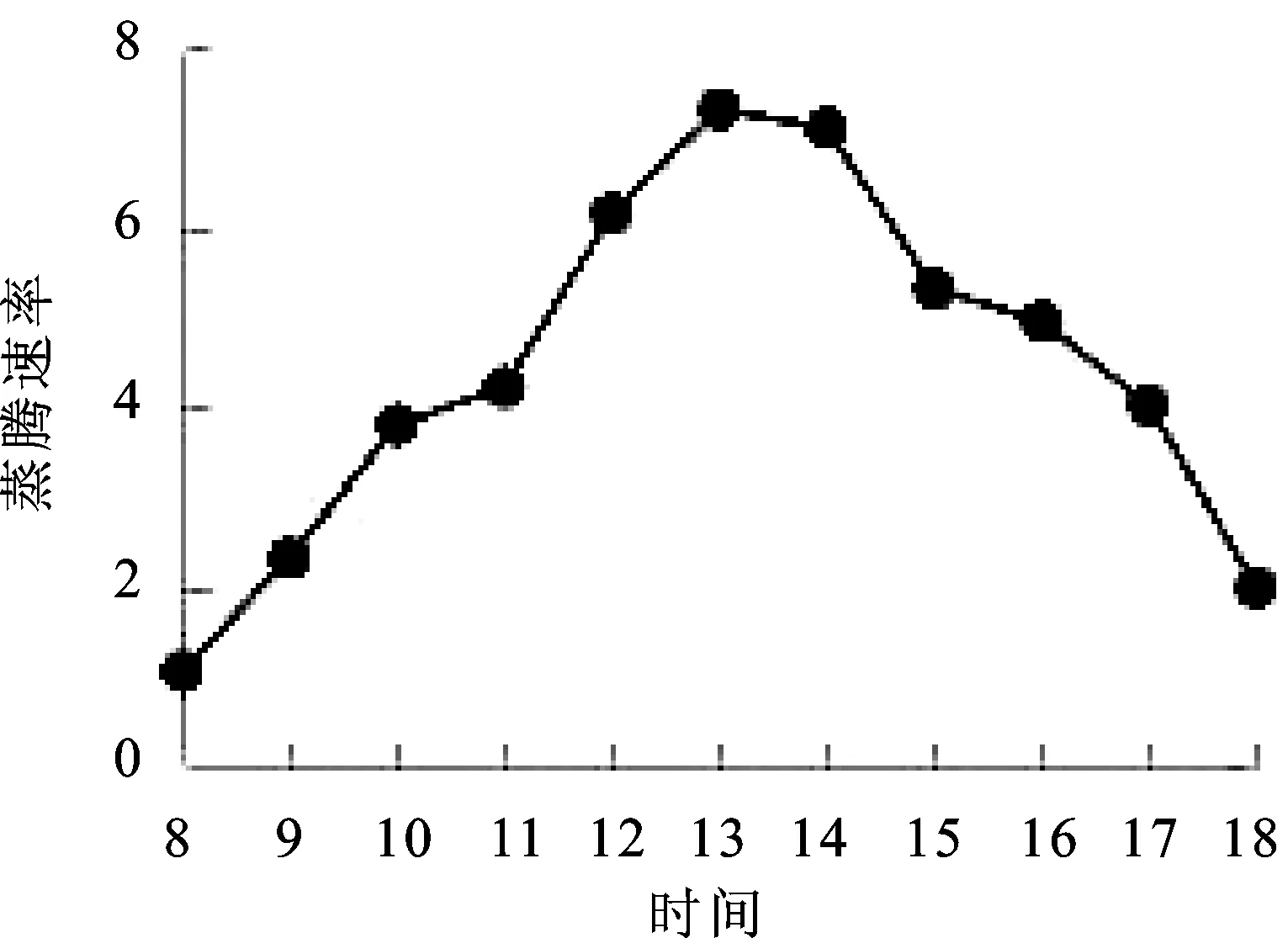
图7 蒸腾速率(Tr)日变化
2.8胞间CO2浓度(Ci)日变化 由图8可知,窝儿七叶片胞间CO2浓度(Ci)日变化呈现“双峰”。8∶00=~9∶00=胞间CO2浓度迅速上升,达到第一个峰值411.62 μmol·m-2·s-1,9∶00=~11∶00=胞间CO2浓度缓慢下降,11∶00=~13∶00=缓慢上升到第二个峰值412.19μmol·m-2·s-1,谷底出现在11∶00=左右,13∶00=~18∶00=缓慢下降到最小值98.43 μmol·m-2·s-1,18∶00=时,胞间CO2浓度数值最低,低于8∶00=的胞间CO2浓度数值,谷底的胞间CO2浓度数值高于8∶00=的胞间CO2浓度数值。胞间CO2浓度是光和生理生态研究中经常用到的一个重要参数,特别是在光合作用气孔限制分析中,Ci的变化方向是确定光合速率变化是否为气孔因素的的主要原因[14]。
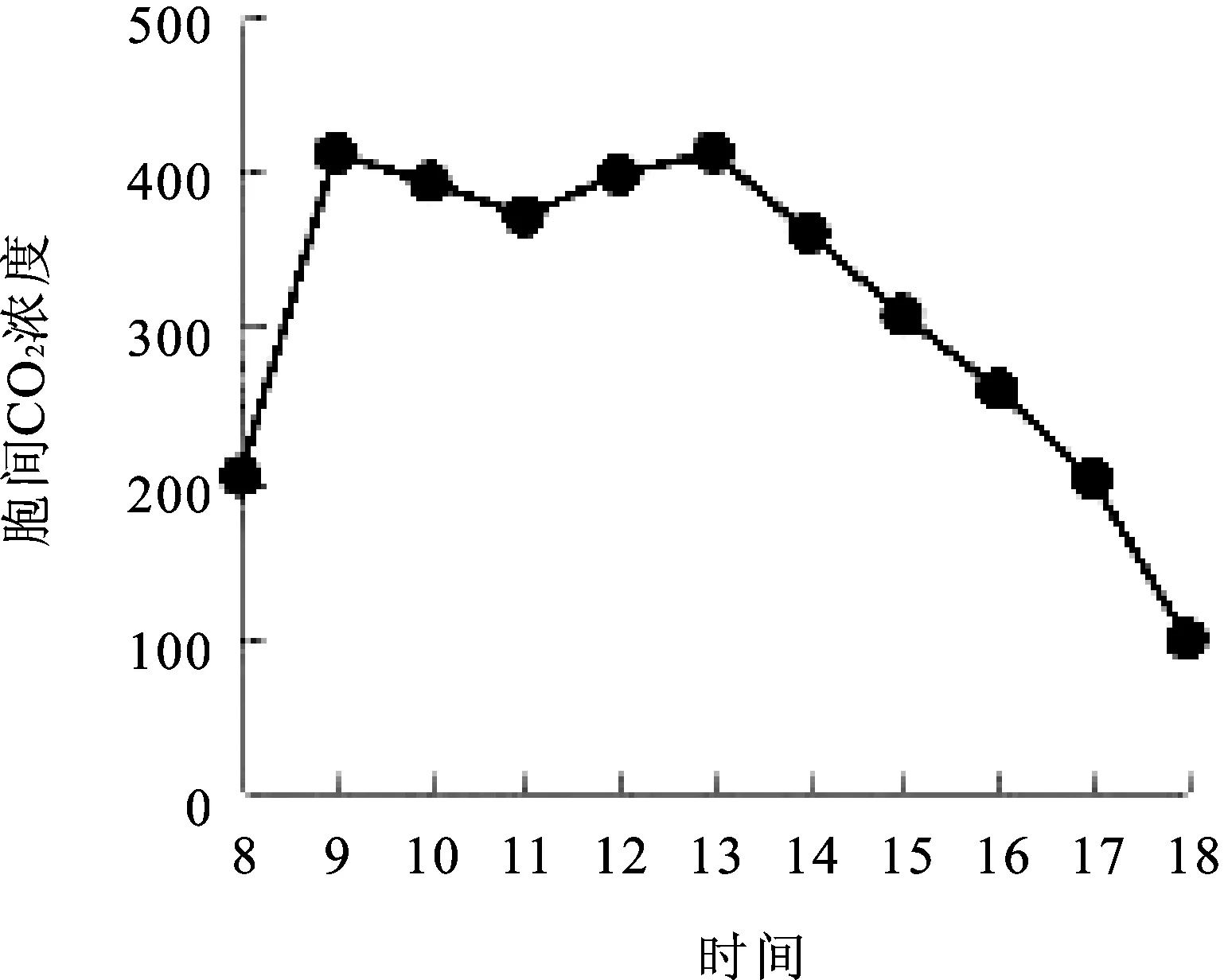
图8 胞间CO2浓度(Ci)日变化
2.9水分利用率(WUE)日变化 由图9可知,窝儿七叶片水分利用率WUE在8∶00=最大,其WUE数值为10.00 μmol·mol-1,8∶00=~10∶00=水分利用率随着时间的延长而逐渐降低,10∶00=~11∶00=水分利用率有缓慢上升的趋势并达到第一个峰值3.75 μmol·mol-1,11∶00=~14∶00=水分利用率又开始缓慢降低保持相对稳定,14∶00=~15∶00=水分利用率缓慢上升并达到第二个峰值2.80 μmol·mol-1,15∶00=~17∶00=缓慢下降保持相对稳定,17∶00=~18∶00=迅速上升并达到最大值4.00 μmol·mol-1,明显低于8∶00=时水分利用率。有研究表明,空气湿度通过影响蒸腾速率来影响水分利用率,空气湿度越低,水分利用率越高[15]。
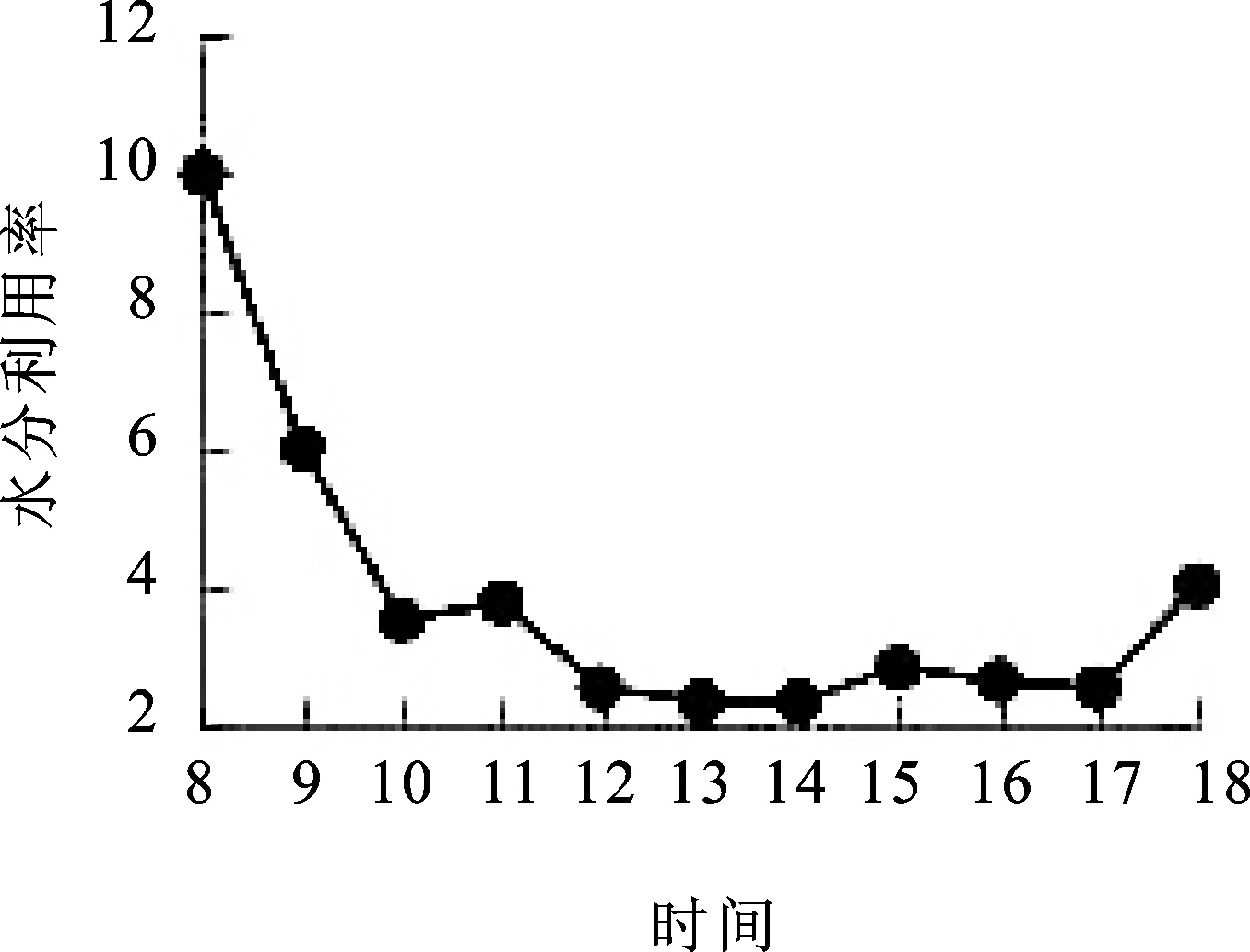
图9 水分利用率(WUE)日变化
2.10气孔限制(Ls)日变化 在水分胁迫条件下,由于气孔导度的限制,导致胞间CO2浓度不能满足光合作用的需求[16]。窝儿七叶片气孔限制(Ls)日变化规律如图10,呈现“双峰”曲线,8∶00=~9∶00=气孔限制迅速下降,9∶00=~12∶00=气孔限制缓慢下降保持稳定,于12∶00=达到最小值,其Ls数值为0.01,12∶00=~18∶00=气孔限制开始上升,于18∶00=达到最大值0.69,18∶00=的气孔限制值明显大于8∶00=的气孔限制值。
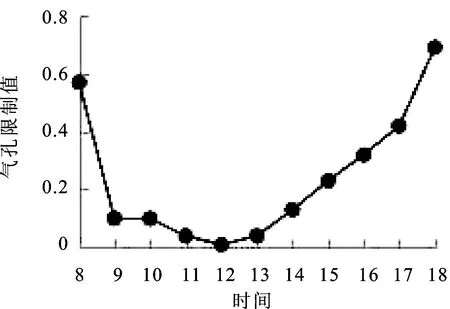
图10 气孔限制(Ls)日变化
2.11窝儿七叶片净光合速率与影响因子的相关性分析 生理生态因子共同作用会引起植物的净光合速率(Pn)的影响,通过对窝儿七叶片净光合速率及其主要生理生态因子进行多元相关性分析(表1),通过Pn与各影响因子间的数值相关分析可知,窝儿七叶片净光合速率数值日变化相关性分析,与Gs呈极显著正相关(r=0.749,P<0.01),与Ci呈极显著正相关(r=0.860,P<0.01),与Tr呈极显著正相关(r=0.817,P<0.01),PAR呈极显著正相关(r=0.932,P<0.01),与RH呈显著负相关(r=-0.628,P<0.05),与Ta呈极显著正相关(r=0.782,P<0.01),与Ls呈极显著负相关(r=-0.898,P<0.01)。分析结果表明,Pn与主要影响因子PAR,Tr,Gs,Ci,呈正相关,与Ca、RH呈负相关。比较各影响因子与窝儿七叶片净光合速率Pn的相关性大小排序为:PAR>Ci>Tr>Ta>Gs>Ca>WUE>RH>Ls,表明光合有效辐射、胞间CO2浓度、蒸腾速率、大气温度、气孔导度是影响窝儿七叶片净光合速率Pn的主要影响因子。
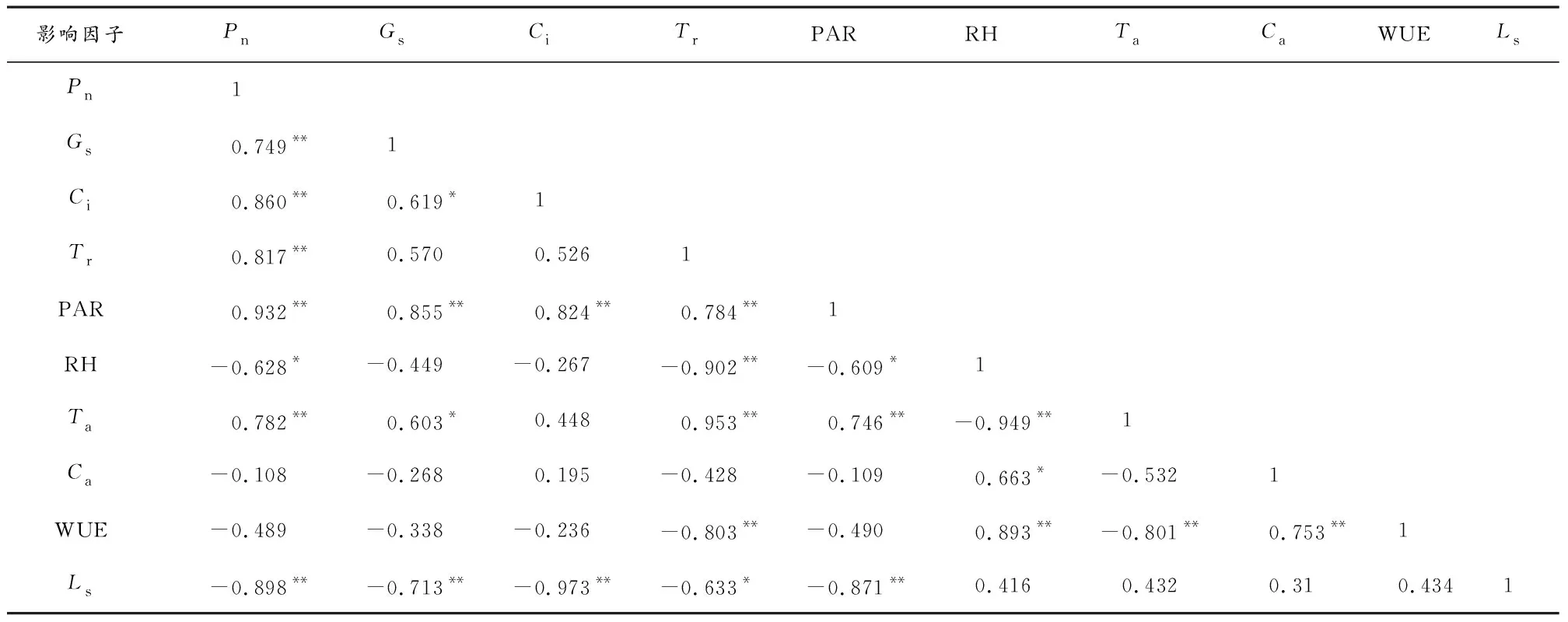
表1 窝儿七叶片净光合速率与影响因子的相关分析
2.12窝儿七叶片的光响应 窝儿七的光合-光响应曲线为图11,Pn最大时其数值为Pmax21.42 μmol·m-2·s-1,此时的光照强度PAR数值为2000 μmol·m-2·s-1,但在所测的光强范围内,窝儿七并未出现光饱和点。当光照强度PAR为零时净光合速率Pn为-2.24 μmol·m-2·s-1,即为光补偿点LCP,此时是窝儿七叶片进行暗呼吸作用。在0~400 μmol·m-2·s-1时,窝儿七叶片的净光合速率Pn随着光照强度PAR的增强而呈显著上升趋势,400~1200 μmol·m-2·s-1时,窝儿七叶片的净光合速率Pn随着PAR的增强而缓慢上升,1200~2000 μmol·m-2·s-1时,净光合速率Pn变化较小,趋于稳定。
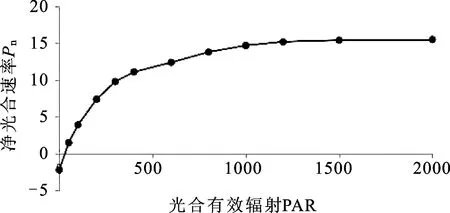
图11 窝儿七叶片光合速率―光合有效辐射响应曲线
3 讨论
光合作用是绿色植物重要的生理特征,太阳是绿色植物光合作用所需能量的直接来源,适宜的光照可以保证植物高产、稳产,遮阴对植物的光合作用、形态结构、生理特性等都有显著影响[17]。本研究结果表明,窝儿七叶片的净光合速率呈现“单峰”形式,存在微弱的“光和午休”现象,这与窝儿七的生存环境有直接的关系。窝儿七为林下植物,由图2,3,5,6,7可知,在8∶00=~11∶00=,随着光有效辐射和温度的上升,气孔导度逐渐增大,蒸腾速率上升,空气相对湿度逐渐变小,净光合速率不断上升,说明温度上升有利于光合速率的增加,这与徐振锋[18]等人的研究相一致。净光合速率和光合有效辐射强度的日变化均为先升后降的趋势,这是因为光有效辐射强度改变引起温度、空气相对湿度等环境因子的变化,而空气相对湿度可以影响气孔导度,并控制CO2摄入量,进一步影响植物的光合速率[19]。11∶00=~13∶00=,随着温度和光有效辐射继续增加,蒸腾速率也在升高,空气相对湿度保持恒定,气孔导度开始缓慢下降,净光合速率逐渐保持相对稳定。大气CO2浓度的变化趋于稳定。通过对窝儿七叶片净光合速率与影响因子的相关分析,得出光合有效辐射、胞间CO2浓度、蒸腾速率、大气温度、气孔导度是影响窝儿七叶片净光合速率Pn的主要影响因子,Pn与Gs呈极显著正相关,与RH呈负相关,说明空气相对湿度大会抑制气孔开闭,减少光合作用。窝儿七叶片在其适宜光合作用时,气孔打开,气孔导度变大,当相对湿度逐渐饱和,温度升高,不宜光合作用时,气孔逐渐闭合,气孔导度变小,说明窝儿七是气孔限制型植物。
本文采用便携式光合测定仪对窝儿七叶片进行光合参数日变化的测定,揭示环境因子对窝儿七叶片光合特性的日变化规律,为选择合理的栽培生境和适宜的种植条件提供了有力的依据,也为提高窝儿七药材品质奠定基础。
