不同磷水平下CO2浓度升高对番茄光合特性和抗氧化酶活性的影响
2022-06-24王月王鲁鑫李浩天刘兴斌韩晓日
王月 王鲁鑫 李浩天 刘兴斌 韩晓日

摘要:通过水培试验研究在低磷(2 μmol/L)、磷充足(2 mmol/L)条件下,大气中不同CO2浓度[(400±50)、(800±50) μmol/mol]对番茄光合特性和抗氧化酶活性的影响。结果表明,磷充足条件下,CO2浓度升高可以显著促进番茄叶片光合速率的提高;而低磷抑制了这种作用。磷充足时,CO2浓度升高显著增加了叶绿素含量,并且叶绿素b含量的增幅明显大于叶绿素a含量;而低磷条件下,CO2浓度升高显著降低了叶绿素含量。与磷充足相比,低磷条件下,番茄叶片的超氧化物歧化酶、过氧化物酶、过氧化氢酶活性明显降低,丙二醛含量升高。但CO2浓度升高明显促进了3种抗氧化酶的活性,并且磷充足条件下促进作用更为显著,同时降低了MDA的含量。因此CO2浓度升高条件下,磷素充足供应可以促进CO2浓度升高产生的正效应。
关键词:番茄;CO2浓度升高;低磷;光合作用;抗氧化酶活性
中图分类号:S145.3;S641.206 文献标志码: A
文章编号:1002-1302(2022)12-0163-05
收稿日期:2021-08-29
基金项目:国家自然科学基金(编号:41602363)。
作者简介:王 月(1977—),女,辽宁北镇人,博士,讲师,主要从事植物营养施肥与土壤肥力研究。E-mail:wangyue1028@163.com。
设施农业中通常施用CO2来补充密闭条件下CO2的缺乏问题,从而增加蔬菜产量和提高品质。CO2施肥对作物的影响还由于全球大气变化而受到更广泛的关注。由于森林砍伐、化石燃料的燃烧等人为活动导致大气中CO2浓度逐年升高,产业革命以前大气中CO2浓度约是280 μmol/mol,目前,大气中CO2浓度已经超过了400 μmol/mol,预计2100年将会达到730~1 020 μmol/mol[1]。
CO2作为植物光合作用的重要原料,其浓度升高势必会对植物光合作用产生影响[2]。有关CO2浓度升高对植物的影响国内外学者进行了大量探索[3-5]。普遍认为在响应短期高浓度CO2时,多数植物会增加净光合作用和减少蒸腾作用[6]。在光合作用中叶绿素发挥着重要作用,它的含量会直接影响植物的光合效率。关于CO2浓度升高对植物叶绿素影响的结论并不一致。研究认为,CO2浓度升高对植物叶绿素含量的影响,表现为促进效应[7]、抑制效应[8]和没有影响[9]。这可能与植物种类以及CO2处理时间等因素不同有关。植物叶片的衰老与光合性能密切相关,因为叶片衰老会影响光合面积以及光合速率。膜质过氧化最重要的产物之一丙二醛(MDA)以及抗氧化酶超氧化物歧化酶(SOD)、过氧化物酶(POD)是标志衰老性状较为常用的敏感指标。通常CO2浓度升高能提高植物的抗氧化能力[10-11],也有研究认为,CO2浓度升高会降低植物的抗氧化酶活性[12]。但以上的研究大多是在单一CO2浓度升高条件下进行的,而CO2浓度升高对植物的影响还与其他环境因子有关[13],其中养分有效性是重要的因素之一,因其在植物响应高浓度CO2时发挥着调控作用[14-15]。磷素作为植物必需的大量营养元素,其有效性直接影响植物光合作用对CO2的响应[16-17]。本研究在不同供磷条件下,探索CO2浓度升高对番茄光合作用和抗氧化系统生理指标的影响,对评价和预测CO2浓度升高条件下番茄植株生长和适应性有一定的实践指导意义,同时为植物的磷素营养管理提供参考依据。
1 材料与方法
1.1 试验材料与方法
试验于2020年11月在沈阳农业大学植物营养学科人工气候室内进行。供试作物为番茄(302番茄)。先将种子消毒催芽,然后播在清洗干净的石英沙中。待幼苗长出2张真叶时进行移栽,将4株长势一致的幼苗转移到1 L的培养器皿中,内装1/2浓度Hoagland营养液,共移栽12盆,供后续进行的4个处理,重复3次使用。培养7 d后,进行不同CO2浓度与供磷量处理。每天光照12 h,光照度是10 000 lx;白天、晚上的温度分别为25、18 ℃。处理后16 d进行相关性状测定。
1.2 供磷水平和CO2浓度
供磷水平设低磷、磷充足2个水平。磷充足营养液配方:4 mmol/L Ca(NO3)2·4H2O,6 mmol/L KNO3,2 mmol/L NH4H2PO4,2 mmol/L MgSO4·7H2O;微量元素的组成同Arnon营养液。低磷营养液配方:NH4H2PO4的浓度为2 μmol/L,以(NH4)2SO4补充铵态氮,其他同磷充足营养液。营养液pH值调至6.0,每3 d換1次营养液。
CO2浓度处理在培养箱(Conviron E7/2,加拿大)中进行,对照CO2处理浓度为(400±50) μmol/mol,升高CO2处理浓度为(800±50) μmol/mol。
1.3 测定项目与方法
1.3.1 番茄叶片光合作用参数测定 每个处理选择具有代表性的番茄植株3株,选取倒数第3张功能叶,利用便携式Ciras-2 型光合仪进行光合参数的测定。包括净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr),并通过公式计算水分利用效率(WUE)=Pn/Tr。测定时使用内置红蓝光源,叶室温度设定为25 ℃。
1.3.2 番茄叶片叶绿素含量测定 每个处理选择具有代表性的番茄植株3株,将主茎上倒数第3张功能叶摘下,放入自封袋中并置冰盒内带回实验室,立即测定叶绿素a、叶绿素b含量[18],并计算总叶绿素含量及叶绿素a含量/叶绿素b含量。
1.3.3 叶片抗氧化酶活性及丙二醛含量的测定 选择番茄主茎上倒数第3张完全展开叶,放入自封袋中并置冰盒内带回实验室,立即进行3种抗氧化酶即超氧化物歧化酶(SOD)[19]、过氧化物酶(POD)[20]、过氧化氢酶(CAT)[21]等的活性以及丙二醛含量[22]的测定。
1.4 数据统计分析
利用SPSS 16. 0 和Excel 分析试验数据并作图。
2 结果与分析
2.1 不同供磷水平下CO2浓度升高对番茄叶片光合作用的影响
由图1可知,无论低磷还是磷充足条件下,CO2浓度升高均显著增加了番茄的光合速率,低磷条件下增加13.0%,磷充足条件下增加26.1%;2个CO2水平下, 均为磷充足处理光合速率显著高于低磷处理。气孔导度以及蒸腾速率在响应CO2浓度升高时均显著降低,低磷和磷充足条件下分别降低69.1%和13.8%、53.9%和21.2%;CO2浓度升高明显促进了番茄叶片的水分利用效率,在低磷和磷充足条件下分别增加145.1%、60.1%。
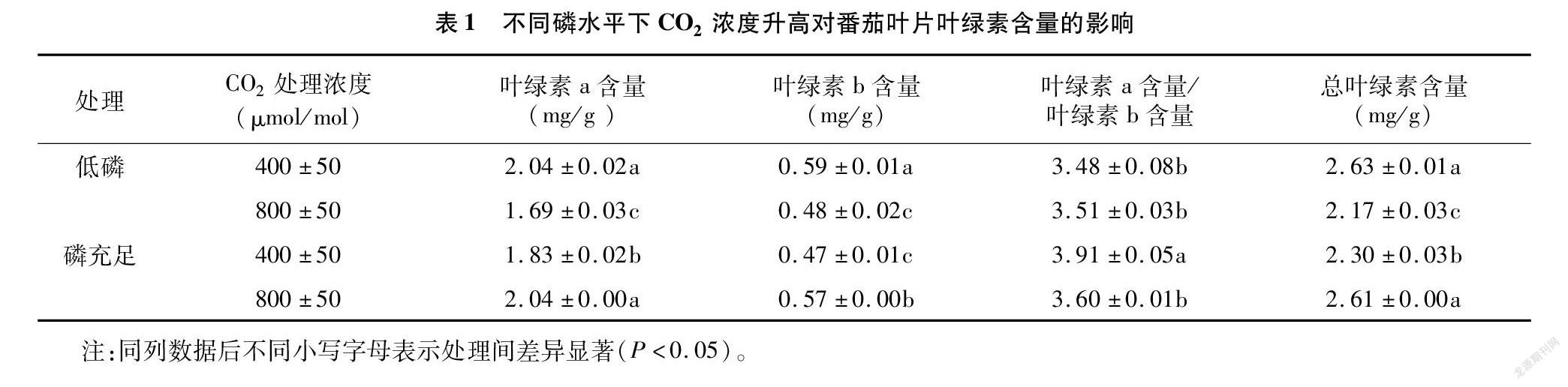
2.2 不同供磷水平下CO2濃度升高对番茄叶片叶绿素含量的影响
从表1可以看出,在低磷条件下,叶片中叶绿素a、叶绿素b和总叶绿素含量在响应CO2浓度升高时显著减少,分别比对照降低了17.2%、18.1%、17.5%,叶绿素a含量/叶绿素b含量的变化不显著;磷充足时,叶绿素a、叶绿素b和总叶绿素含量在响应CO2浓度升高时均显著提高,分别比对照提高了11.5%、21.0%、13.5%,其中叶绿素a含量和叶绿素b含量的比值显著降低。
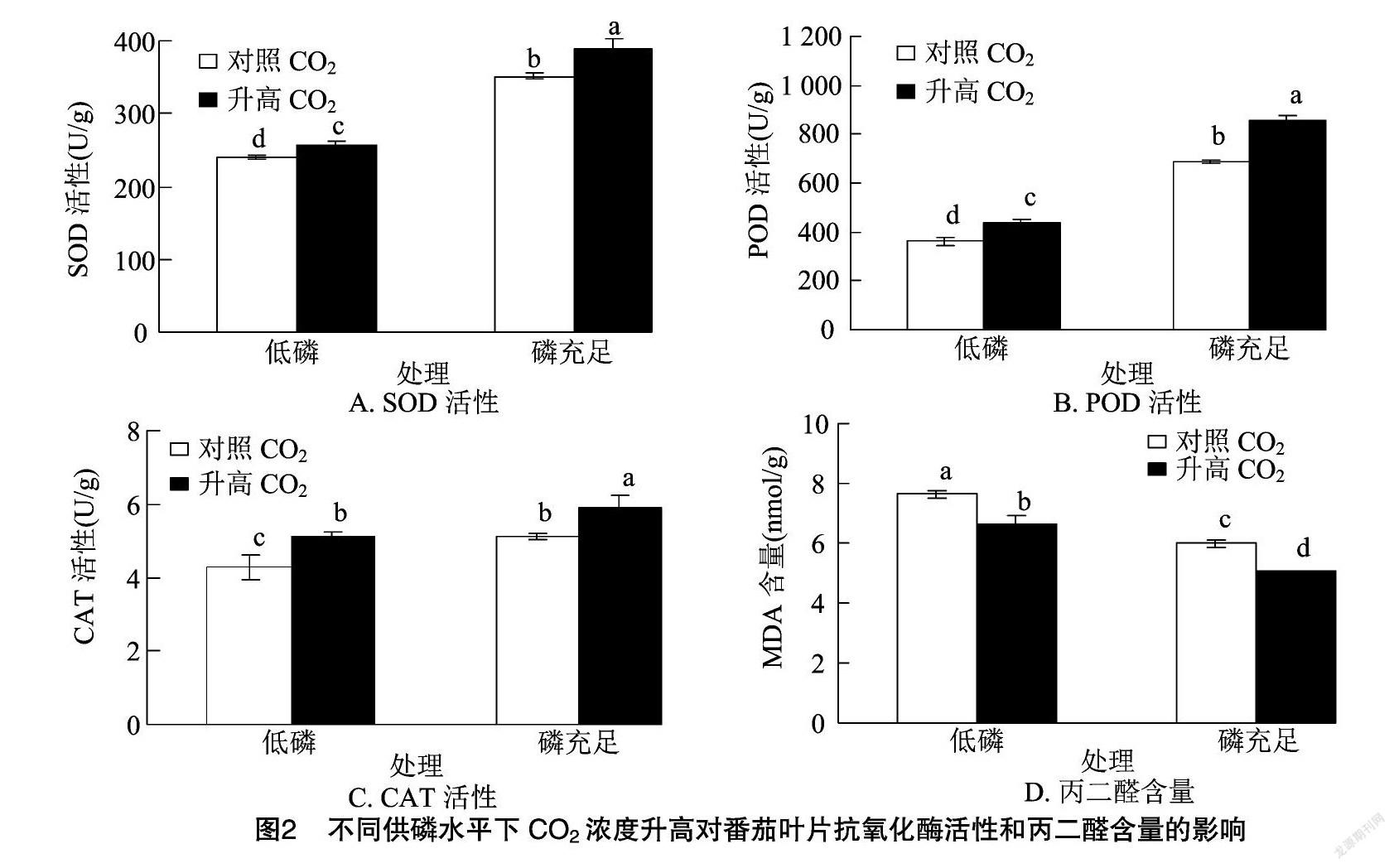
2.3 CO2浓度升高对番茄叶片抗氧化酶活性和丙二醛含量的影响
对照CO2条件下,与磷充足相比,低磷使番茄叶片SOD、POD和CAT的活性分别降低了31.9%、47.3%、16.2%(图2-A、图2-B、图2-C),而MDA的含量比磷充足条件下增加了27.5%(图2-D);CO2浓度升高后,SOD、POD和CAT的活性分别增加了7.7%、21.2%、19.3%,同时MDA的含量降低了13.3%。磷充足条件下,番茄叶片中SOD、POD和CAT的活性在响应CO2浓度升高时,分别比低碳处理增加10.6%、25.1%、15.8%,MDA的含量降低了15.4%,且处理间差异显著。
3 讨论与结论
众多研究表明,CO2浓度升高通常会刺激C3植物的光合作用和生长潜力[23-24]。本研究结果表明,无论在低磷还是在磷充足条件下,CO2浓度升高均促进了番茄叶片的净光合速率,但低磷限制了这种效应,这与相关研究结果[25]一致,表明磷供应不足会使大中CO2浓度升高,对植物生长的促进作用减弱。无论CO2浓度高或低,低磷均显著降低了番茄的光合速率。低磷条件下植物净光合速率降低并非气孔导度降低减少了CO2的供应,而是由于非气孔因素阻碍了对CO2的利用[26],主要是因为磷不足影响了植物的代谢过程[27]。高浓度CO2条件下植物叶片的气孔往往会关闭[28],减少植物与大气之间的气体交换,从而使气孔导度下降,蒸腾速率降低[29],这在本研究中进一步得到了证实。由于光合速率增加和蒸腾速率下降,本研究中番茄水分利用效率显著升高,这与相关研究结果[30-31]一致。
相关研究表明,CO2浓度升高对植物叶片中叶绿素含量,叶绿素a含量和叶绿素b含量的比值均有影响,但结论不一致。长期生长在CO2浓度升高条件下的植物,其叶片中叶绿素的含量会降低,叶绿素a含量和叶绿素b含量的比值会下降[32-33]。本研究中在对照CO2条件下,低磷处理的叶绿素含量显著高于磷充足处理,可能是由于缺磷时植物生长缓慢,干物质积累少,所以叶绿素含量相对较高;低磷条件下,当CO2浓度升高时叶绿素含量显著降低,可能是由于CO2浓度升高促进了其光合作用,积累了更多的干物质,所以叶绿素含量降低。当磷充足时,CO2浓度升高使叶绿素含量增加,并且叶绿素a含量与叶绿素b含量的比值降低,说明磷充足时CO2浓度升高有利于叶绿素的形成并且更有利于形成叶绿素b,这或许是CO2浓度升高增强植物光合作用的原因之一,因为叶绿素b含量增加可使补光色素蛋白复合体的生成增多,从而增强了叶绿体对光能的吸收。也有研究认为,CO2浓度升高导致叶绿素a含量/叶绿素b含量升高[34-35]或者不变[36],这可能与植物种类或生育时期等不同有关。
MDA的积累量间接反映了植物受胁迫的程度[37-38]。SOD、POD、CAT这3种抗氧化保护酶,可以清除植物体内的活性氧等超氧化物,防止膜脂过氧化,提高植物的生理活性、减缓其衰老[39]。本研究中低磷导致了番茄叶片中MDA的含量显著升高,这与在甘蔗以及在水稻叶片上的研究结果[40-41]一致,因此低磷导致了超氧自由基的增多并造成了膜脂过氧化程度的提高。同时缺磷处理番茄叶片SOD、POD、CAT等3种抗氧化酶活性显著降低,但是CO2浓度升高增加了3种抗氧化酶的活性,因此增强了抗氧化能力,使其膜脂过氧化程度减弱,进而降低了番茄叶片中的MDA含量,表明在一定期间内高浓度CO2对保护番茄叶片防止氧化损伤起着重要的作用。这与在樱桃番茄以及在番茄幼苗上的研究结果[42-43]类似。而彭长连等发现,CO2浓度升高条件下水稻的氧化损伤程度降低,同时几种抗氧化酶的活性也降低,认为高浓度CO2条件下氧化损伤程度的降低主要是由于生成的活性氧减少所致[12]。林久生等发现,在小麦的整个生长期几种抗氧化酶的活性均表现为CO2倍增条件下明显高于CK,并且H2O2含量也高于CK,认为CO2倍增条件下小麦氧化损伤程度的减轻并不是由于活性氧含量的下降,而主要是由于细胞内抗氧化能力的增强[44]。可见由于试验使用材料的种类、发育阶段、CO2浓度和处理时间以及其他生长环境等不同,使得高浓度CO2对植物自由基的产生以及清除的影响试验结果不尽一致。
综上所述,CO2浓度升高增强了番茄的光合效率,而低磷抑制了这种作用;磷充足条件下CO2浓度升高有利于番茄叶片叶绿素的形成,并且更有利于叶绿素b的形成,同时也增强了番茄抗氧化系统的保护能力,进而促进光合作用。因此CO2浓度升高条件下,磷素的充足供应可以促进CO2浓度升高产生的正效应。
参考文献:
[1]Kumar A,Nayak A K,Sah R P,et al. Effects of elevated CO2 concentration on water productivity and antioxidant enzyme activities of rice (Oryza sativa L.) under water deficit stress[J]. Field Crops Research,2017,212:61-72.
[2]Redda A R,Rasinen G K,Raghawendra A S. The impact of global elevated CO2 concentration on photosynthesis and plant productivity[J]. Current Science,2010,99(1):46-57.
[3]DeLucia E H,Hamilton J G,Naidu S L,et al. Net primary production of a forest ecosystem with experimental CO2 enrichment[J]. Science,1999,284(5417):1177-1179.
[4]郝兴宇,韩 雪,李 萍,等. 大气CO2浓度升高对绿豆叶片光合作用及叶绿素荧光参数的影响[J]. 应用生态学报,201 2(10):2776-2780.
[5]刘 超,胡正华,陈 健,等. 不同CO2浓度升高水平对水稻光合特性的影响[J]. 生态环境学报,2018,27(2):246-254.
[6]Long S P,Ainsworth E A,Rogers A,et al. Rising atmospheric carbon dioxide:plants FACE the future[J]. Annual Review of Plant Biology,2004,55:591-628.
[7]Pinter Jr P J,Idso S B,Hendrix D L,et al. Effect of free-air CO2 enrichment on the chlorophyll content of cotton leaves[J]. Agricultural and Forest Meteorology,1994,70(1/2/3/4):163-169.
[8]Rao M V,Hale B A,Ormrod D P. Amelioration of ozone-induced oxidative damage in wheat plants grown under high carbon dioxide (role of antioxidant enzymes)[J]. Plant Physiology,1995,109(2):421-432.
[9]Reeves D W,Rogers H H,Prior S A,et al. Elevated atmospheric carbon dioxide effects on sorghum and soybean nutrient status[J]. Journal of Plant Nutrition,1994,17(11):1939-1954.
[10]莊明浩,李迎春,郭子武,等. 大气CO2浓度升高对毛竹叶片膜脂过氧化和抗氧化系统的影响[J]. 生态学杂志,2012,31(5):1064-1069.
[11]刘 筱. 模拟CO2浓度升高和降水改变对薄荷生理生态特性的影响[D]. 贵阳:贵州大学,2020.
[12]彭长连,林植芳,林桂珠. 加富CO2条件下水稻叶片抗氧化能力的变化[J]. 作物学报,1999,25(1):39-43.
[13]赵天宏,王美玉,张巍巍,等. 大气CO2浓度升高对植物光合作用的影响[J]. 生态环境,2006,15(5):1096-1100.
[14]Lloyd J,Farquhar G D. The CO2 dependence of photosynthesis,plant growth responses to elevated atmospheric CO2 concentrations and their interaction with soil nutrient status. Ⅰ. General principles and forest ecosystems[J]. Functional Ecology,1996,10(1):4.
[15]牛晓光,杨荣全,李 明,等. 大气CO2浓度升高与氮肥互作对玉米光合特性及产量的影响[J]. 中国生态农业学报,2020,28(2):255-264.
[16]Conroy J P,Smillie R M,Küppers M,et al. Chlorophyll a fluorescence and photosynthetic and growth responses of Pinus radiata to phosphorus deficiency,drought stress,and high CO2[J]. Plant Physiology,1986,81(2):423-429.
[17]Lewis J D,Griffin K L,Thomas R B,et al. Phosphorus supply affects the photosynthetic capacity of loblolly pine grown in elevated carbon dioxide[J]. Tree Physiology,1994,14(11):1229-1244.
[18]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000.
[19]Giannopolitis C N,Ries S K.Superoxide dismutases:Ⅱ. purification and quantitative relationship with water-soluble protein in seedlings[J]. Plant physiology,1997,59(2):315-318.
[20]陈贻竹,B.帕特森. 低温对植物叶片中超氧物歧化酶、过氧化氢酶和过氧化氢水平的影响[J]. 植物生理学报,1988,14(4):323-328.
[21]Cakmak I,Marschner H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase,ascorbate peroxidase,and glutathione reductase in bean leaves[J]. Plant Physiology,1992,98(4):1222-1227.
[22]Heath R L,Packer L.Photoperoxidation in isolated chloroplasts:Ⅰ. Kinetics and stoichiometry of fatty acid peroxidation[J]. Archives of Biochemistry and Biophysics,1968,125(1):189-198.
[23]Kant S,Seneweera S,Rodin J,et al. Improving yield potential in crops under elevated CO2:integrating the photosynthetic and nitrogen utilization efficiencies[J]. Frontiers in Plant Science,2012,3:162.
[24]Morales A,Yin X Y,Harbinson J,et al. In silico analysis of the regulation of the photosynthetic electron transport chain in C3 plants[J]. Plant Physiology,2017,176(2):1247-1261.
[25]Conroy J P. Influence of elevated atmospheric CO2 concentrations on plant nutrition[J]. Australian Journal of Botany,1992,40(5):445.
[26]朱隆靜,喻景权. 不同供磷水平对番茄生长和光合作用的影响[J]. 浙江农业学报,2005,17(3):120-122.
[27]Jacob J,Lawlor D W. Stomatal and mesophyll limitations of photosynthesis in phosphate deficient sunflower,maize and wheat plants[J]. Journal of Experimental Botany,199 42(8):1003-1011.
[28]Mott K A. Sensing of atmospheric CO2 by plants[J]. Plant,Cell and Environment,1990,13(7):731-737.
[29]Centritto M,Lucas M E,Jarvis P G. Gas exchange,biomass,whole-plant water-use efficiency and water uptake of peach (Prunus persica) seedlings in response to elevated carbon dioxide concentration and water availability[J]. Tree Physiology,2002,22(10):699-706.
[30]Klaiber J,Najar-Rodriguez A J,Piskorski R,et al. Plant acclimation to elevated CO2 affects important plant functional traits,and concomitantly reduces plant colonization rates by an herbivorous insect[J]. Planta,2013,237(1):29-42.
[31]Brestic M,Zivcak M,Hauptvogel P,et al. Wheat plant selection for high yields entailed improvement of leaf anatomical and biochemical traits including tolerance to non-optimal temperature conditions[J]. Photosynthesis Research,2018,136(2):245-255.
[32]Oberbauer S F,Strain B R,Fetcher N. Effect of CO2-enrichnient on seedling physiology and growth of two tropical tree species[J]. Physiologia Plantarum,1985,65(4):352-356.
[33]Socias F X,Medrano H,Sharkey T D. Feedback limitation of photosynthesis of Phaseolus vulgaris L grown in elevated CO2[J]. Plant,Cell and Environment,1993,16(1):81-86.
[34]Nie G Y,Long S P,Garcia R L,et al. Effects of free-air CO2 enrichment on the development of the photosynthetic apparatus in wheat,as indicated by changes in leaf proteins[J]. Plant,Cell and Environment,1995,18(8):855-864.
[35]卢从明,张其德,冯丽洁,等. CO2浓度倍增对谷子拔节期和灌浆期光合色素含量和PSⅡ功能的影响[J]. 植物学报,1997,39(9):874-878.
[36]韦彩妙,林植芳,孔国辉.提高CO2浓度对两种亚热带树苗光合作用的影响[J]. 植物学报,1996,38(2):123-130.
[37]高 婷,张 杰,马瑞红,等. NaCl胁迫对黑籽南瓜生长和生理特性的影响[J]. 江苏农业科学,2020,48(6):122-124,137.
[38]赵 嫚,陈仕勇,李亚萍,等. 外源GABA对盐胁迫下金花菜种子萌发及幼苗抗氧化能力的影响[J]. 江苏农业学报,202 37(2):310-316.
[39]高小丽,孙健敏,高金锋,等. 不同基因型绿豆叶片衰老与活性氧代谢研究[J]. 中国农业科学,2008,41(9):2873-2880.
[40]万美亮,邝炎华,陈建勋. 缺磷胁迫对甘蔗膜脂过氧化及保护酶系统活性的影响[J]. 华南农业大学学报,1999,20(2):1-6.
[41]潘晓华,刘水英,李 锋,等. 低磷胁迫对不同水稻品种叶片膜脂过氧化及保护酶活性的影响[J]. 中国水稻科学,2003,17(1):57-60.
[42]Karim M F,Hao P F,Nordin N H B,et al. CO2 enrichment using CRAM fermentation improves growth,physiological traits and yield of cherry tomato (Solanum lycopersicum L.)[J]. Saudi Journal of Biological Sciences,2020,27(4):1041-1048.
[43]Zhang Y,Yao Q,Shi Y,et al. Elevated CO2 improves antioxidant capacity,ion homeostasis,and polyamine metabolism in tomato seedlings under Ca(NO3)2-induced salt stress[J]. Scientia Horticulturae,2020,273:109644.
[44]林久生,王根軒. CO2 倍增对渗透胁迫下小麦叶片抗氧化酶类及细胞程序性死亡的影响[J]. 植物生理学报,2000,26(5):453-457.
