浙南人工红树林秋茄叶片光合作用与荧光特性动态研究
2022-06-23周成黄颖杨华蕾杜明卉张颖唐剑武
周成 黄颖 杨华蕾 杜明卉 张颖 唐剑武

关键词:红树林秋茄;动态变化;光合作用;叶绿素荧光;相关性
0引言
滨海湿地(红树林、盐沼、海草)生态系统因其生物量以及土壤的强大碳汇能力而越来越多地被认为是重要的碳库[1],科学家把这类碳库所储存的碳定义为滨海湿地“蓝碳”[2-3].近年来,随着对全球生态系统研究的逐渐深入,科学家研究发现,在单位面积上滨海湿地生态系统的碳汇能力高于多数森林生态系统[1,4-6],有着巨大的碳汇潜力.红树林生态系统是滨海湿地生态系统中碳汇能力较强的生态系统之一,能够提供优质的底栖生物居所,也有着巨大生态效益[5].而植物的光合作用是滨海湿地生态系统固碳的关键过程之一[7-8].已有研究表明,植物的光合作用和叶绿素荧光动态变化受外界光照和温度等环境因子影响.例如,余文英等[9]研究表明,辽河三角洲湿地的芦苇叶片的净光合速率、气孔导度、蒸腾速率等光合参数一般在中午前后会达到最高,且在生长季中夏季最高、秋季最低;徐晓迪等[10]研究表明,闽楠的光合参数以及荧光参数日变化存在差异,其日变化特征主要呈倒U型与双峰状;黄丽等[11]研究发现,秋茄、桐花树、白骨壤和木榄4种红树林的光合参数在夏季达到最高,而在冬季达到最低;陈敬华等[12]研究发现,湿地松与马尾松叶片实际光化学效率的日变化呈U型,而非光化学淬灭与之相反,表明强光下植物为了保护自身会把过剩的光能以热能形式释放到外界;洪文君等[13]研究表明,海南假韶子的净光合速率、蒸腾速率与叶绿素荧光参数均表现为7月大于11月,并揭示水分是造成该现象的关键因子,等等.因此,不同植被叶片光合作用均有着自身的规律,探明叶片光合作用动态规律,利于从叶片尺度上了解滨海湿地碳通量的动态过程,为今后从叶片尺度上定量估算垂向固定的碳提供科学依据.
秋茄作为我国最耐寒的红树林物种,近期被逐渐引种到浙江温州,以此来替换与控制互花米草[14].但是,高纬度种植的秋茄叶片光合生理特性以及光合参数与荧光参数间的关系研究較少.因此,本文将以浙江温州市平阳县鳌江口红树林秋茄为研究对象,探究其叶片光合作用与荧光特性的日变化、月变化,以及它们之间的关系,为近期秋茄北移引种工程和红树林生态系统碳通量评估提供科学依据.
1材料与实验方法
1.1 研究区概况
浙南蓝碳生态过程监测实验站(120.57°E,27.58°N)位于浙江省温州市平阳县鳌江口红树林秋茄种植区.属于中亚热带海洋性季风气候区,全年光照充足、气候湿润,年平均气温为15~22℃,最高月平均气温为31℃,最低月平均气温为5℃,降雨主要集中在3—9月.全年都受到潮汐、地表径流等共同影响.
1.2 研究材料
在浙南蓝碳生态过程监测试验站附近四年生的人工红树林内,选取3株红树植物秋茄冠层上部、生长良好、完全展开的阳性叶子作为测量对象.
1.3 气体交换参数测量
选择晴朗天气,分别于2019年4月26日(4月)、6月11日(6月)、7月24日(7月)、8月26日(8月)、9月(取8月与10月的平均值代替9月)、10月30日(10月)、12月2日(代替11月)、12月28日(12月)开始进行2~3d数据监测,利用LI-6800F便携式光合-荧光自动测量系统测量气体交换参数.仪器参数设置为:流速500L/min,其他环境因子为环境值,测定时间为8:00—16:00.每小时测量3株秋茄,每株选取2~3枚叶片,重复记录2~3个数据值,取其平均值作为该时刻的气体交换参数测定值.测量的参数包括净光合速率(A)、蒸腾速率(E)、气孔导度(Gsw)、胞间浓度(Ci)、光合有效辐射(PAR)、空气温度(T)以及湿度(RH%).计算叶片水利用效率(WUE),即A/E.
1.4 叶绿素荧光参数测量
在进行气体交换测量的过程中,利用LI-6800F便携式光合-荧光自动测量系统荧光叶室同步记录叶片叶绿素荧光参数测量值,后期取其平均值作为该时刻的叶绿素荧光参数测量值.测量的参数包括实际光化学量子效率(фPSⅡ)、电子传递效率(ETR)、非光化学淬灭系数(NPQ)、光适应下的最大光化学量子效率(/)以及光化学淬灭系数(qP).
1.5 数据处理
采用MicrosoftExcel2019进行数据统计分析和绘图,相关性分析采用SPSS21.0软件进行.
2结果与分析
2.1 不同月份叶片光合作用的日变化
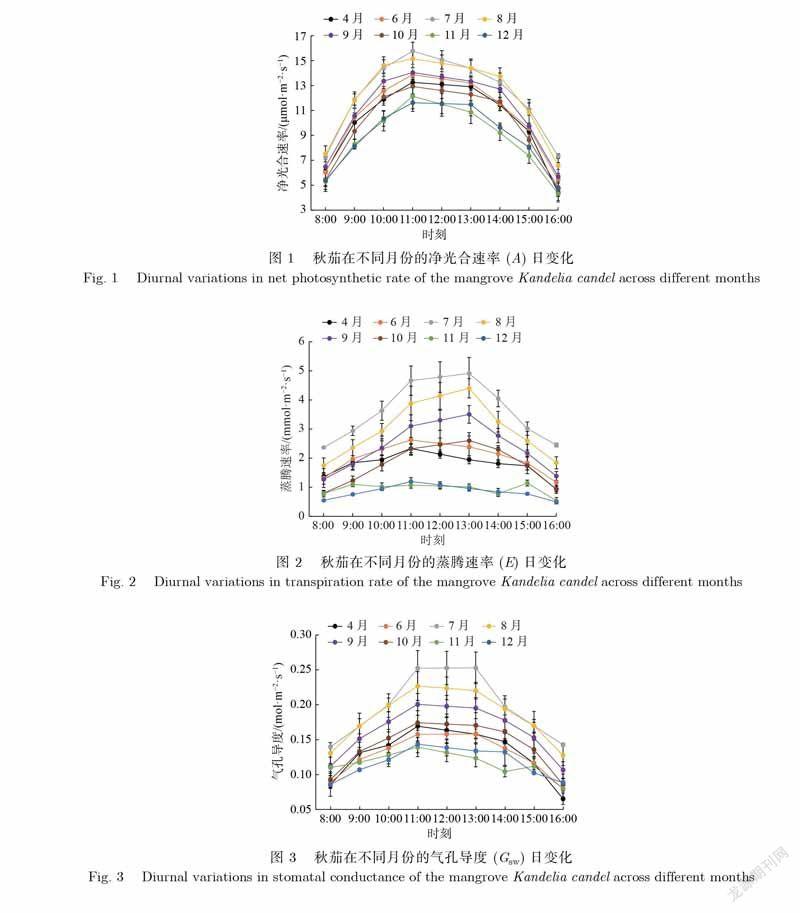
净光合速率(A)的大小直接反映了植物在当前生长环境中的光合能力.如图1所示,秋茄在不同月份的A日变化无明显差异,在各个月份均呈明显的倒U型,且在11时前后出现峰值.各月份峰值最大出现在7月,为15.8μmol·m–2·s–1,最小出现在12月,为11.5μmol·m–2·s–1.可能的原因是7月的温度、光照等因子比12月的更加适宜秋茄生长,促使植物出现较高的净光合速率.秋茄的日均A值在7月、8月最大,分别为12.3、12.2μmol·m–2·s–1,而在11月、12月最小,分别为8.8、9.0μmol·m–2·s–1.7月、8月较高的温度和光照促使秋茄维持着较高的光合能力,而11月、12月的温度较低且光照较弱,抑制了其光合作用,同时在一定程度上说明了7月、8月是秋茄的生长季.
蒸腾速率(E)反映着植物向外界散失水分的多少.如图2所示,秋茄在不同月份的E日变化差异显著.7月、8月、9月、10月4个月份的秋茄E日变化均呈倒U型,峰值出现在13时前后.4月、6月、12月3个月份的也呈倒U型,而峰值出现在11时前后,低于7月、8月、9月的峰值.但是,11月的日变化呈双峰状,出现两个峰值,分别在9时与15时前后,均低于其他月份的峰值.秋茄的日均E值由大到小依次为7月、8月、9月、6月、10月、4月、11月、12月,对应值分别为3.65、3.02、2.41、2.03、1.8、1.78、0.94、0.84mmol·m-2·s-1.由此可见,秋茄的E值从4月开始逐渐增加,到7月达到最大值,之后逐渐减小,到12月达到最小值.
气孔导度(Gsw)反映着植物的气孔张开程度,影响着植物的光合能力.如图3所示,秋茄在不同月份的Gsw日变化差异显著.8月、9月、10月、12月4个月份的Gsw日变化均呈倒U型,峰值均出现在中午11时前后,明显低于7月的最大值.而6月、7月、10月3个月份的Gsw均是上午先增大,中午11—13时基本不变,下午开始减小.11月的Gsw与E日变化类似,呈双峰状,但两个峰值出现的时间并不完全一致,分别在11时前后和15时前后.秋茄的Gsw日均值由大到小依次为7月、8月、9月、10月、4月、6月、12月、11月,对应值分别为0.197、0.185、0.163、0.142、0.131、0.128、0.117、0.116mol·m–2·s–1.由此可见,各月份的气孔导度有着显著差异,在夏季的7月、8月最高,而在接近冬季的11月、12月最低.
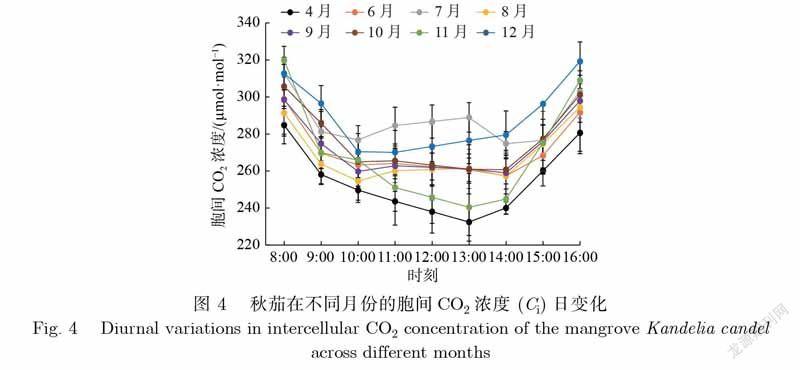
胞间CO2浓度(Ci)表示植物叶片细胞间的CO2浓度的大小,影响着植物的光合能力.如图4所示,秋茄在不同月份的Ci日变化差异显著.除7月份外,其他月份的日变化均呈U型,谷值出现的时间不一致,但均在10—14时.在各月份上均表现为早上和晚上的Ci值高,中午的Ci值较低,原因可能是中午的叶片细胞在高光照下光合能力强,同化CO2的量大,致使Ci值较小.秋茄Ci日均值由大到小依次为12月、7月、10月、9月、6月、11月、8月、4月,对应值分别为288.3、287.3、276.2、272.4、270.7、269.1、268.7、254.1μmol·mol–1.由此可见,各月份的Ci值变化表现为从4月开始逐渐增大,7月达到最大,之后开始减小,11月达到最小,12月又增大,说明温度变化影响着秋茄的Ci值.
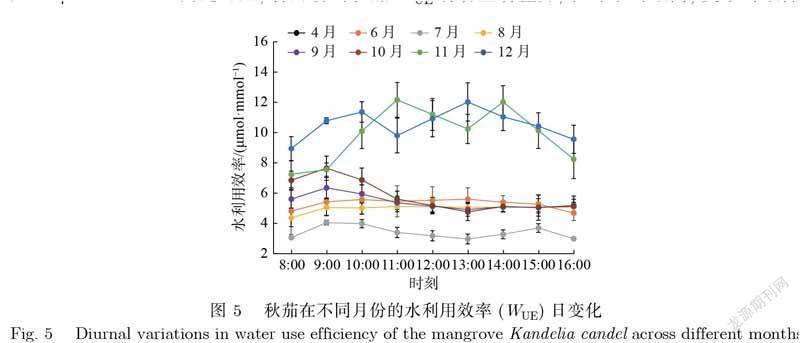
水利用效率(WUE)反映着植物在散失一定水分下同化CO2的基本效率.如图5,秋茄在不同月份的WUE日变化差异显著.4月、7月、11月、12月4个月份的WUE日变化均呈明显的双峰状,但峰值出现并不完全一致,4月、7月、12月的WUE峰值分別在10时和13时前后,而11月的WUE峰值分别在11时和14时前后.9月和10月的WUE日变化表现为上午的值比下午的大,且上午呈倒U型,峰值均出现在9时前后.但是,6月与8月的WUE日变化不显著.秋茄的WUE日均值由大到小依次为12月、11月、10月、4月、9月、6月、8月、7月,对应值分别为10.54、9.88、5.81、5.77、5.40、5.31、5.00、3.40μmol·mmol–1.由此可见,各月份的秋茄WUE有着显著差异,在冬季时最高,夏季时最低.
2.2 不同月份叶片叶绿素荧光的日变化
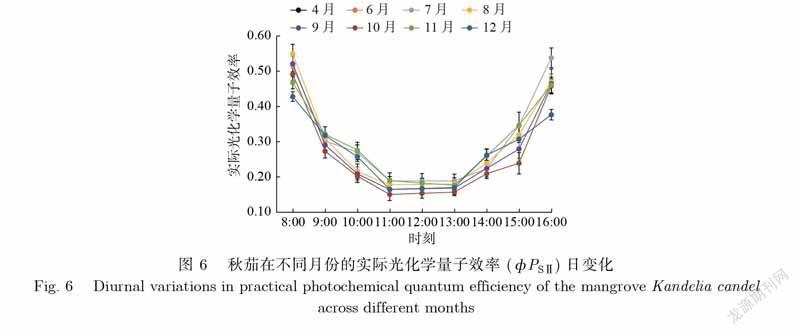
植物光合系统Ⅱ实际光化学反应量子效率(фPSⅡ)反映了植物光合系统Ⅱ中叶绿素将所吸收的光能用于光化学反应的比例,也是衡量植物光合能力的重要参数.如图6所示,秋茄在各月份的фPSⅡ日变化差异显著,均呈U型,表现为上午фPSⅡ值开始降低,中午12时前后最低,下午又开始增加,其主要驱动因子可能是光照.秋茄的фPSⅡ日均值由大到小依次为7月、8月、11月、6月、9月、12月、4月、10月,对应值分别为0.306、0.293、0.289、0.285、0.276、0.272、0.263、0.259.由此可见,秋茄的фPSⅡ日均值月变化明显,夏季7月、8月的值较其他月份大,说明在夏季秋茄叶片传递电子通量大,有着更高的光合能力.
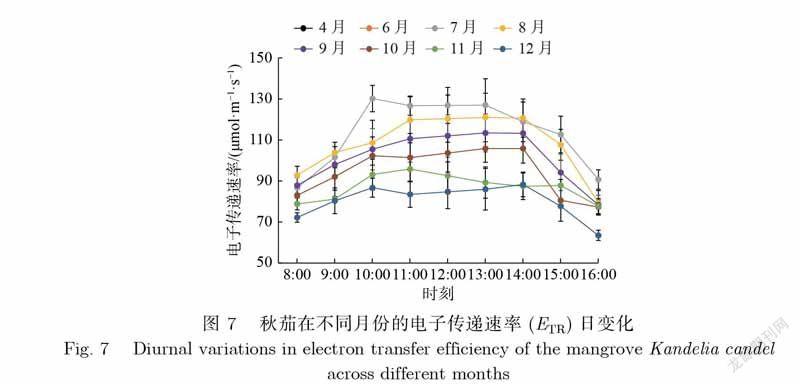
叶片电子传递速率(ETR)代表植物叶片在不同的光照强度下光合系统Ⅱ传递光合电子的相对速率.如图7,秋茄在不同月份的ETR日变化差异显著,均呈倒U型,与фPSⅡ日变化相反.具体表现为ETR值上午逐渐增大,中午12时前后变化幅度较小,下午开始逐渐降低.秋茄的ETR日均值由大到小依次为7月、8月、6月、9月、4月、10月、11月、12月,对应值分别为113.4、108.2、105.0、101.4、96.6、94.6、87.0、80.3μmol·m–1·s–1.由此可见,ETR日均值月变化呈明显差异,夏季7月、8月最高,冬季11月、12月最低,说明秋茄叶片光合系统Ⅱ在夏季时传递光合电子的能力较强,而在冬季时较弱.
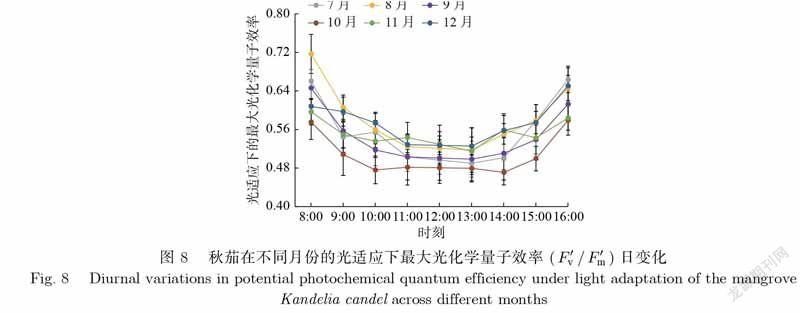
植物光合系统Ⅱ的最大光化学反应量子效率(/)代表植物叶片在光适应下,光合系统Ⅱ进行光化学反应时利用光能的最大比例,是衡量植物潜在光合能力的指标.如图8所示,秋茄在不同月份的/日变化差异显著,均呈U型,表现为早晚高、中午低,且在中午前后变化幅度较小.秋茄的F′vF′mF′vF′m/日均值由大到小依次为8月、12月、7月、11月、9月、6月、10月、4月,对应值分别为0.58、0.57、0.55、0.55、0.54、0.53、0.51、0.50.由此可见,秋茄的/日均值月变化不明显,在夏季7月、8月和冬季11月、12月都是比较高的,但冬季的实际光合能力低于夏季的,说明植物在冬季由于受低温等因子影响而出现光抑制现象.
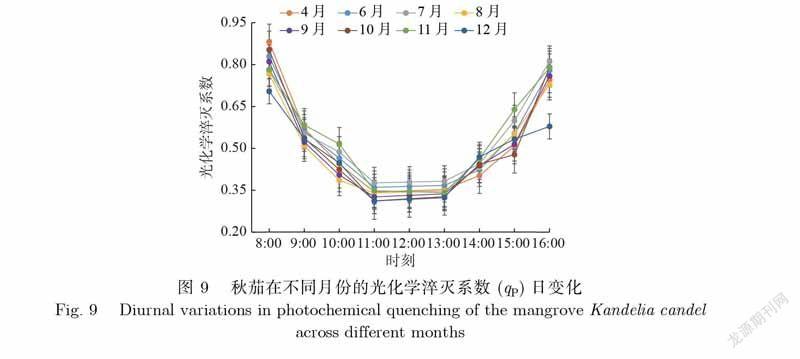
光化学淬灭系数(qP)与фPSⅡ类似.如图9所示,秋茄在不同月份的qP日变化差异显著,均呈U型,与фPSⅡ日变化类似,具体表现为qP值上午逐渐降低,中午变化幅度较小,下午又开始逐渐增大.秋茄的qP日均值由大到小依次为7月、11月、6月、4月、10月、8月、9月、12月,对应值分别为0.53、0.53、0.52、0.51、0.50、0.49、0.49、0.47.由此可见,秋茄的qP日均值月变化不明显,且各月份的qP日均值均约为0.5,说明秋茄在不同月份均有着较好的光合活性.
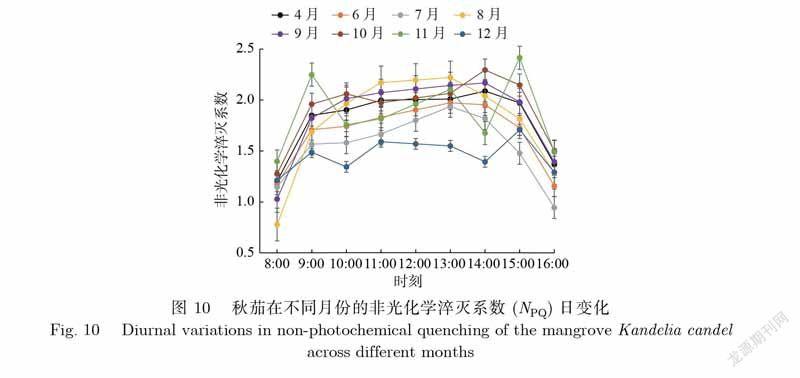
非光化学淬灭系数(NPQ)反映植物叶片所吸收的光能用于热能的大小变化,是衡量植物热耗散能力的重要指标.如图10所示,秋茄在不同月份的NPQ日变化差异显著.7—10月均呈倒U型;4月、6月均呈双峰型;11月、12月均出现了3个峰值,变化起伏较多,且峰值的出现时间并不完全一致.秋茄的NPQ日均值由大到小依次为10月、11月、9月、4月、8月、6月、7月、12月,对应值分别为1.92、1.87、1.86、1.82、1.80、1.69、1.55、1.46.由此可见,秋茄的NPQ日均值月变化显著,在秋季的9月、10月和11月明显大于其他月份,说明该时期内植物对光能的利用率较低.
2.3 叶片光合因子及叶绿素荧光因子的相关性
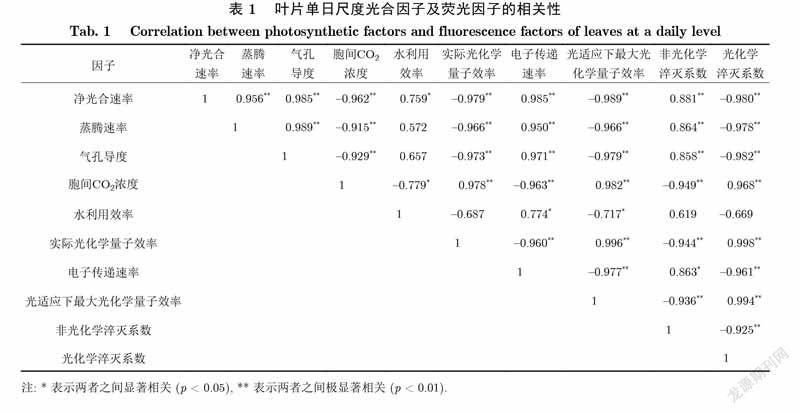
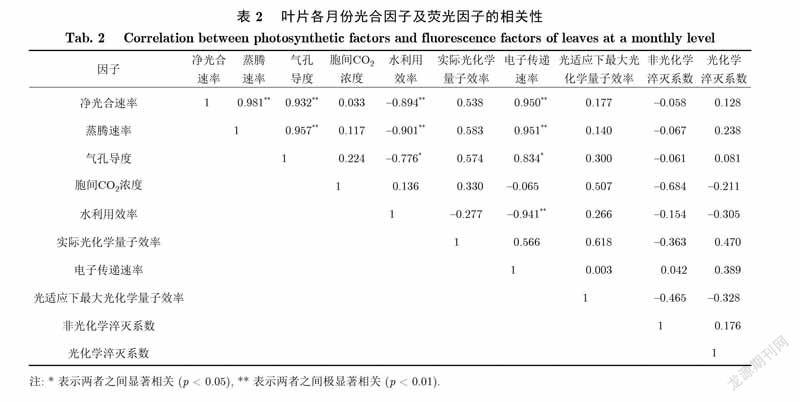
如表1所示,秋茄叶片的单日尺度光合因子及叶绿素荧光因子相关性分析结果表明,各因子之间密切相关.光合因子之间的关系是:秋茄叶片的A、E、Gsw两两之间呈极显著正相关,与Ci均呈极显著负相关;WUE与A呈显著正相关,而与Ci呈显著负相关.荧光因子之间的关系是:秋茄叶片的фPSⅡ、/、qP两两之间呈极显著正相关,分别与ETR、NPQ呈极显著负相关.光合因子与荧光因子之间的关系是:秋茄叶片的A、E、Gsw分别与ETR、NPQ呈极显著正相关,而与фPSⅡ、/、qP均呈极显著负相关;Ci与фPSⅡ、/、qP均呈极显著正相关,与ETR、NPQ均呈极显著负相关;WUE与ETR呈显著正相关,而与/呈显著负相关.
如表2所示,秋茄叶片各月份的光合因子及叶绿素荧光因子相关性分析结果表明,各因子之间的关系较日尺度的弱.光合因子之间的关系是:秋茄叶片的A、E、Gsw两两之间呈极显著正相关;WUE与A、E呈极显著负相关,与Gsw呈显著负相关.荧光因子之間关系不显著,仅фPSⅡ与/的相关系数大于0.6,呈正相关.光合因子与荧光因子之间的关系是:秋茄叶片的A、E分别与ETR呈极显著正相关,Gsw与ETR呈显著正相关,WUE与ETR呈极显著负相关.
3讨论
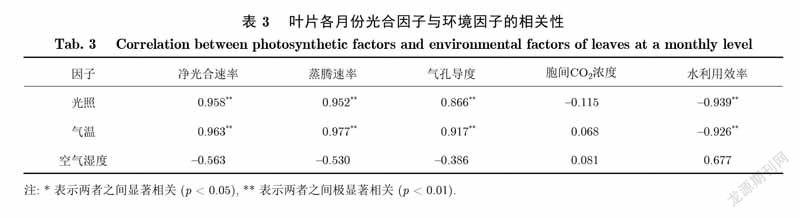
不同月份植被所处的环境因子差异会导致不同月份光合作用的动态变化及其差别[15],如表3所示.本研究发现,各月份的秋茄在叶片尺度上光合参数日变化主要呈U型(如A、Gsw、E)、倒U型(如Ci).秋茄7月、8月份的各时刻A值均高于其他几个月份.一方面是因为夏季优越的光照与温度条件比较适合这时环境中的秋茄叶片光合作用的需求.另一方面是因为叶片在7月前还处于逐渐苏醒阶段,各机能尚未完全活跃,因而叶片光合能力较弱,而处于7月、8月的叶片已经完全苏醒,光合能力达到最强,同时外界也存在着优越的环境条件,内外因素一同造成该时期叶片具有高光合能力.相反,11月、12月除8时与16时外各时刻A值均明显低于其他几个月份,原因可能是该时期的低温以及低光照等外界环境促使秋茄出现光抑制现象,而且秋茄叶片也由于这些外界条件的改变而降低了自生的叶绿素,使得秋茄叶片光合能力变弱[16].另外,各月份各光合参数的日变化存在差异,说明导致秋茄光合能力变化的重要因子是外界环境因子.除11月、12月外,当11时前Gsw值逐渐达到最大时,A值随之增大,而Ci值随之下降,说明气孔张开程度是导致秋茄叶片光合速率与胞间浓度变化的主要原因.研究还发现,叶片WUE的大小受所处环境因子的影响,即WUE与环境因子PAR、T呈极显著负相关,而与环境空气湿度呈正相关,但不显著,这与刘宪锋等[17]及史晓亮等[18]的研究结果类似.因此,秋茄11月、12月WUE的值明显高于其他月份,7月WUE值最低.尽管秋茄叶片在夏季7月、8月有着高净光合速率,但水利用效率却比较低,可能是因为秋茄在夏季有着较高的气孔张开程度,使得蒸腾速率较高,从而WUE值会出现未增反减的现象[19].
植物的叶绿素荧光是研究植物光合作用的一种敏感探针[20-21].本研究发现,各叶绿素荧光参数的日变化主要呈U型(如фPSⅡ、qP、/)、倒U型(如ETR)、双峰型(如NPQ).在日内光照变化的影响下,秋茄的фPSⅡ、/、qP在中午光照最强时达到最低,而NPQ除7、8月外在此时均未达到最高,却在14、15时达到最大.表明在强光照下,秋茄光合系统Ⅱ中叶绿素将所吸收的光能用于光化学反应的比例降低,光合系统Ⅱ的光合活性降低,同时在植物自我保护机制下光合系统内部将过剩的光能转成热能释放到外界环境,从而NPQ在午间有个较高的值[22].下午时фPSⅡ、/、qP又逐渐恢复到之前的水平,说明植物的光合活性受到抑制后是可逆的,以及该保护机制十分有效地保护了植物的光合系统.此外,ETR在午间能够维持着较高的值,也进一步说明,植物的自我保护机制为能够维持较高的光合系统Ⅱ传递光合电子的能力提供了基础,保证植物在高光下有着较高的光合能力.研究还发现,12月的фPSⅡ均值接近其他月的,同时12月的NPQ均值最低,说明在低温低光照的12月,秋茄叶片会降低吸收的光能转为热能的比例来提高实际光化学效率,从而缓解冬季光照相对较弱的影响[12].
植物叶片光合作用与叶绿素荧光联系紧密[21].本研究发现,秋茄叶片在单日尺度上光合因子和荧光因子之间的相关性比较高,除WUE外,均达到0.8以上.佐证了秋茄叶片气孔导度是导致净光合速率日变化和胞间浓度日变化的直接因子,以及фPSⅡ、ETR、NPQ有着紧密联系是午间植物维持较高净光合速率的直接因子.此外,研究分析各月份(日均值)光合因子与荧光因子之间的相关性发现,A、E、Gsw、WUE、ETR相关性较大,均达到0.7以上.佐证了秋茄叶片气孔导度是导致各月份净光合速率变化和蒸腾速率变化的直接因子.同时,A、E均与WUE负相关,达到0.9以上,佐证了叶片的光合速率和蒸腾速率是造成各月份WUE差异的直接因子,即夏季WUE低,而冬季WUE高.
另外,通过对比南方秋茄叶片光合参数还发现[11,23-25](表4),本研究中的浙江温州鳌江口的红树秋茄相比于福建漳江口和泉州湾河口的红树秋茄,其叶片净光合速率提高了1~2μmol·m–2·s–1,最大净光合速率提高了约3μmol·m–2·s–1,水利用效率提高了至少1倍,气孔导度无较大差异.此外,相比福建漳江口的50年生红树秋茄和泉州湾河口的红树秋茄,其蒸腾速率降低了3.33~4.56mmol·m–2·s–1,一定程度上减少了叶片水分的散失;其光饱和点也比较高,约为1500μmol·m–2·s–1,相比福建泉州湾河口的红树秋茄,提高了约500μmol·m–2·s–1.以上说明了,红树秋茄在向北移植至浙江温州鳌江口后,它的光合能力有一定的提高,其生长适应情况是比较好的.
4结论
通过对浙南人工秋茄叶片光合作用与叶绿素荧光不同月份日变化进行研究,本文得出以下重要结论:
(1)各参数日变化特征主要为倒U型和U型.除NPQ外,各参数变化均表现为早晚变化大、中午变化幅度小.夏季7月、8月为秋茄主要生长旺盛季节,其光合能力明显优于其他月份,而冬季12月则相反.
(2)单日上,除WUE外,光合因子与荧光因子的相关性显著;而各月份上,A、E、Gsw、WUE、ETR之间的相关性显著.表明E、Gsw、ETR是影响秋茄叶片A日变化及月变化的重要因子.
(3)光合因子与荧光因子的相关性显著,叶绿素荧光一定程度上可以用来探测植物光合作用的变化情况.
