扑草净胁迫下香根草叶片的代谢组学研究
2022-06-16孙仕仙石傲傲
张 坤 孙仕仙 石傲傲 郑 毅
(1.云南农业大学植物保护学院,云南 昆明 650201;2.西南林业大学湿地学院,云南 昆明 650233;3.云南开放大学,云南 昆明 650223)
扑草净(2,4−双(异丙基氨基)−6−(甲硫基)−s−三嗪)是一种选择内吸传导性三嗪类除草剂,与其他三嗪类除草剂相比,扑草净在三嗪环上用烷硫基取代了第3 个氯原子,这样使扑草净既保留了三嗪类除草剂的特性,如除草的广谱性和长效性,又能降低除草剂对作物的毒害作用[1]。由于此特性使得扑草净在很长一段时间内在国内外被广泛应用,随着扑草净在动植物中不断被检测出有较高残留量的问题,扑草净的安全使用问题及对环境和人体的潜在危害也逐渐被人们所重视。随着研究的不断深入,发现扑草净具有类似苯环的结构,是一种内分泌干扰物质,化学性质较为稳定,可长期存在于环境及生物体中[2]。由于残留扑草净的废水排入河流、湖泊或者海洋,中国黄渤海、海南八门湾、莱州湾等水域均有扑草净被检出,许多水产品也被检出扑草净残留超标,严重影响了中国水产品出口贸易[3−4]。因此,寻找一种经济、环保的去除水体中扑草净污染的方法成为目前环境领域亟待解决的问题,其中近些年被关注的植物修复是生物修复领域的一个热点,其为去除土壤和水体中的污染物质提供了一种新的可能性。与物理化学修复相比,植物修复成本更低,更能为大众所接受,因为植物修复可利用植物根茎叶对污染物的积累和富集作用,以及根际微环境区域中植物−微生物−土壤的相互作用达到去除污染物的目的,且具有很好的环境生态效益[5−6]。
香根草(Vetiveria zizanioides)是1 种非侵入性且生长迅速的多年生草本植物,可以耐受和积累金属[7−9]。香根草可以耐受不同的水体环境,包括酸性矿山废水等[9−10]。此外,香根草具有大量的自我再生根,可以形成斑块并隔离金属[9]。项目组前期研究发现香根草对于扑草净有较强的耐受和去除能力[11−13],因此探讨香根草对扑草净的耐受机理,有助于将来更深入地将其应用于水体修复领域。而植物体内的代谢物是生物体表型的基础,能更直观有效地了解其生物学过程及其机理[14]。鉴于扑草净对植物的作用机制之一是影响植物的光合作用,为了明确香根草在扑草净胁迫下的代谢策略,本研究以扑草净胁迫下香根草叶片为研究对象,基于对其代谢物的定性定量分析,分析在扑草净刺激后香根草叶片代谢物的应答机制。
1 材料与方法
1.1 材料来源
试验于2020 年5—7 月在西南林业大学格林温室进行,供试药品中的97%扑草净标准品购买于济南仁诺化工有限公司,供试材料香根草(分蘖苗)购买于江西红壤研究所。试验前先将植株用自来水清洗干净,再用高锰酸钾溶液消毒,然后用去离子水清洗3 遍,将其放入盛有1/2 改良Hoagland 营养液的塑料桶中,适应性培养1~2 周,选取长势良好,大小一致的植株,用于试验。
1.2 试验方法
1.2.1 试验设计
将温室养殖的香根草用水培种植,每桶5 L培养液,加入 5 mg/L 未添加抑菌剂的扑草净。分别于试验第5 天(LM−5 d)、第10 天(LM−10 d)采集植物的叶片样品,以未加扑草净处理的香根草叶片为对照(LM−CK),每个处理采集质量为3 g 的香根草叶片,3 次重复,取样后立即加入液氮并放入−80 ℃冰箱中保存,干冰运送至武汉迈维代谢公司进行非靶向代谢组检测。
1.2.2 代谢物定性与定量分析方法
基于迈维代谢公司自建数据库MWDB(metware database),根据二级谱信息进行物质定性,分析时去除了同位素信号,含K+、Na+、NH4+离子的重复信号,以及本身是其他更大分子量物质的碎片离子的重复信号。
代谢物定量是利用三重四级杆质谱的多反应监测模式(MRM)分析完成。在MRM 模式中,四级杆首先筛选目标物质的前体离子(母离子),排除掉其他分子量物质对应的离子以初步排除干扰。获得不同样本的代谢物质谱分析数据后,对所有物质质谱峰进行峰面积积分,并对其中同一代谢物在不同样本中的质谱出峰进行积分校正。
1.3 广泛靶向代谢组数据分析
1.3.1 差异代谢物筛选
差异代谢物筛选标准:1)选取差异倍数值(fold change)≥2 和fold change≤0.5 的 代谢 物,在对照组和实验组中差异为2 倍以上或0.5 以下,则认为差异显著。2)在上述的基础上,选取OPLS−DA 模型的变量重要性投影(VIP)≥1的代谢物。VIP 值表示对应代谢物的组间差异在模型中各组样本分类判别中的影响强度,一般认为VIP≥1 的代谢物则为差异显著。将差异倍数以2 为底取对数的值,分别将上调和下调排在前10 位差异代谢物挑选出来进行进一步分析。
1.3.2 差异代谢物功能分析
利用KEGG 数据库对差异代谢物进行代谢途径的查询,将差异显著代谢物在代谢途径中进行注释。按照注释结果对差异代谢物在KEGG 中进行通路类型分类,并进行KEGG 通路富集。
2 结果与分析
2.1 不同处理时间叶片差异代谢物的特征
2.1.1 差异代谢物数量分析
本试验共测得香根草叶片的差异代谢物分属于黄酮、生物碱、有机酸、酚酸、核苷酸及其衍生物、氨基酸及其衍生物、木质素和香豆素、其他类、鞣质、脂质、萜类等,具体数量分析见表1。由表1 可知:在LM−5 d vs.LM−10 d 组,变化的一共有48 种,下调的共有27 种,上调的有21 种;在LM−CK vs.LM−10 d 组,变化的一共有151 种,下调的有86 种,上调的有65 种;LM−CK vs.LM−5 d组,变化的一共有121 种,下调的有74 种,上调的与47 种。一共有121 种,下调的有74 种,上调的与47 种。

表1 各处理组间的差异代谢物的数量Table 1 Quantity of differential metabolites in different treatments 个
2.1.2 差异代谢物种类分析
香根草不同处理时间下代谢物种类和数量的差异分析见图1。由图1 可知:胁迫处理与对照相比,代谢物下调的数量要高于上调的数量,可能是由于植物受到胁迫后,影响了根系向叶片的养分供应,从而影响了叶片的正常代谢,代谢产物总体数量有所降低。扑草净胁迫5 d 时与对照相比,下调数量最多的代谢物为黄酮,有19 种;其次为生物碱和脂质,分别为14 种和13 种。上调数量最多的是核苷酸及其衍生物,生物碱和其他类(如微生物、糖、醇类)均有7 种,有机酸和氨基酸及其衍生物均有6 种。扑草净胁迫10 d 与5 d 时相比,叶片中的代谢产物数量变化幅度较小,每类物质的上调数量均小于5 种,黄酮和核苷酸及其衍生物均为4 种,脂质和酚酸类均为3 种。下调代谢物数量最多的是氨基酸及其衍生物,为7 种;其次是生物碱,为6 种;其余种类代谢物下调数量均小于5 种。扑草净胁迫10 d 与对照相比,数量差异比较大的是核苷酸及其衍生物和酚酸类,均比对照有所增加;与5 d 相比,黄酮的下调数量有所减少,新增加了1 种萜类物质。
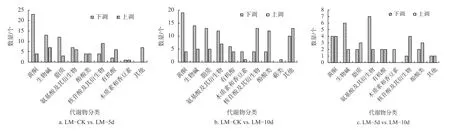
图1 扑草净胁迫下香根草叶片代谢产物差异分析Fig.1 Differential analysis of metabolites in V.zizanioides leaves under prometryn stress
2.2 不同处理时间叶片差异代谢物的KEGG 分析
2.2.1 KEGG 分类分析
在扑草净胁迫下,对不同时间香根草叶片中鉴定的差异代谢物通路注释结果进行比较,结果见图2。
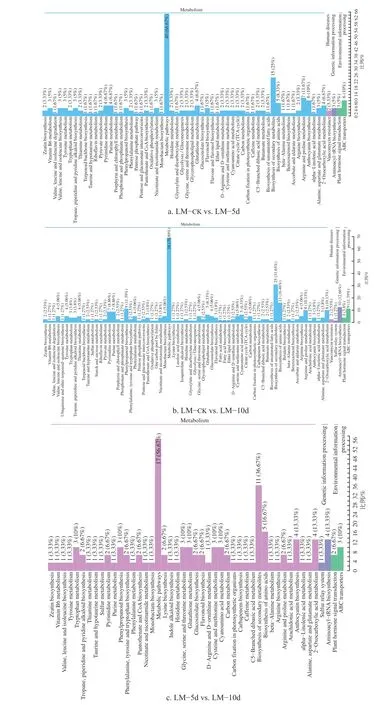
图2 叶片不同处理间差异代谢物的KEGG 分类分析Fig.2 KEGG classification analysis of differential metabolites among different treatments in leaves
扑草净胁迫5 d 时香根草叶片的代谢产物和对照比较结果表明:差异代谢物涉及代谢途径(40 种,占66.67%)、次生代谢产物的生物合成(15 种,占25%)、精氨酸和脯氨酸代谢(7种,占11.67%)、花青素生物合成(6 种,占10%)、氨基酸生物合成(5 种,占8.33%)、2−氧羰基酸代谢(4 种,占6.67%)、嘧啶代谢(4 种,占6.67%)、嘌呤代谢(4 种,占6.67%)、谷胱甘肽代谢(4 种,占6.67%);其次是环境信息处理,涉及ABC 转运体和植物激素信号转导(3 种,5%)。
扑草净胁迫10 d 时香根草叶片的代谢产物和对照比较结果表明:差异代谢物涉及代谢途径(56 种,占70.89%)、次生代谢产物的生物合成(25 种,占31.65%)、氨基酸生物合成(13 种,占16.46%)、类苯基丙烷代谢途径、精氨酸和脯氨酸代谢(8 种,占10.13%);其次是遗传信息处理,其中差异数量最多的是氨基酰−tRNA 生物合成(10 种,占1.66%);最后是环境信息处理(涉及ABC 转运体(9 种,11.39%)和植物激素信号转导(3 种,占3.8%)。
扑草净胁迫10 d 时香根草叶片的代谢产物和5 d 比较结果表明:差异代谢物涉及代谢途径(17 种,占56.67%)、次生代谢产物的生物合成(11 种,占36.67%)、氨基酸生物合成(5 种,占16.67%)、花青素生物合成(4 种,占13.33%)、2−氧羰基酸代谢(4 种,占13.33%);其次是遗传信息处理,其中差异数量最多的是氨基酰−tRNA 生物合成(4 种,占13.33%);最后是环境信息处理,涉及ABC 转运体(3 种,10%)和植物激素信号转导(2 种,占6.67%)。
2.2.2 KEGG 富集分析
对扑草净胁迫下香根草叶片的差异代谢物进行KEGG 富集分析,结果见图3。由图3 可知,胁迫5 d 时与对照相比,选择富集显著且差异代谢物数量较多的2 个通路为:1)花青素生物合成途径,2)精氨酸和脯氨酸代谢途径;选择富集程度较大的且富集显著度也较大的2 个通路为:1)植物激素信号转导途径,2)维生素B6 代谢途径。胁迫10 d 时与对照相比,选择富集显著且差异代谢物数量较多的4 个通路为:1)氨酰-tRNA 生物合成,2)硫代葡萄糖苷生物合成,3)硫胺素代谢,4)精氨酸和脯氨酸代谢途径;选择富集程度及显著度较大的2 个通路为:1)植物激素信号转导途径,2)卟啉和叶绿素代谢。胁迫5 d与胁迫10 d 相比,选择富集显著且差异代谢物数量较多的2 个通路为:1)花青素的生物合成,2)植物激素信号转导途径;选择富集程度较大且富集显著度的1 个通路为:花生四烯酸代谢。

图3 叶片不同处理间差异代谢物的KEGG 富集分析Fig.3 KEGG enrichment analysis of differential metabolites among different treatments in leaves
2.3 不同处理时间叶片高差异倍数代谢物分析
将叶片中的组间代谢物按差异倍数排序,把上调前10 位和下调前10 位的代谢物挑选出来,结果见图4。
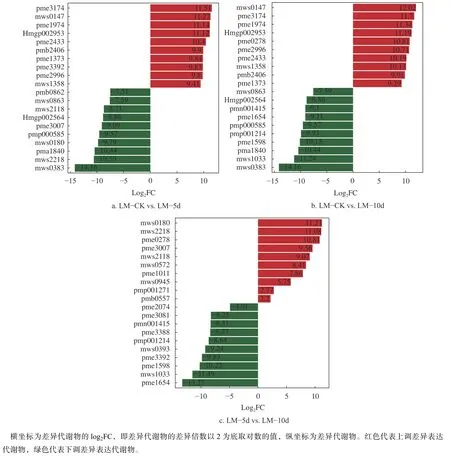
图4 叶片不同处理间上、下调差异倍数排名前10 位的代谢物Fig.4 The top 10 metabolites in the fold difference of up- and down-regulation among different treatments in leaves
由图4 可知,胁迫5 d 时与对照相比,上调代谢物差异倍数从高到低的是胞苷−5'−单磷酸、β−羟基异戊酸、丙酮酸、苜蓿素−O−葡萄糖醛酸−O−鼠李糖苷、二乙醇胺、溶血磷脂酰胆碱 17:0、2'−脱氧胞苷−5'−单磷酸、天竺葵素 3−O−葡萄糖苷、4−羟基香豆素和儿茶酚;下调代谢物差异倍数从高到低的是顺−10−十七碳烯酸、咖啡碱、对香豆酰胆碱、龙胆酸、芹菜素−7−O−(6−O−丙二酰基葡萄糖苷)、尿苷 5'−二磷酸、苜蓿素−7−O−二葡萄糖醛酸苷、根皮苷、2−脱氧核糖−5−磷酸和溶血磷脂酰胆碱 12:0。其中上调的代谢物中,丙酮酸属于有机酸,参与精氨酸和脯氨酸、酪氨酸、苯丙氨酸、丙酮酸、丁酸、烟酸和烟酰胺代谢等多个通路;二乙醇胺参与甘油磷脂代谢通路。下调的代谢物中,咖啡碱属于生物碱,参与咖啡因代谢通路;龙胆酸属于酚酸类,参与酪氨酸代谢通路;根皮苷属于黄酮类,参与黄酮类生物合成通路。
胁迫10 d 时与对照相比:上调代谢物差异倍数从高到低的是β−羟基异戊酸、胞苷−5'−单磷酸、丙酮酸、苜蓿素−O−葡萄糖醛酸−O−鼠李糖、2,6−二氨基庚二酸、4−羟基香豆素、二乙醇胺、儿茶酚、溶血磷脂酰胆碱 17:0、2'−脱氧胞苷−5'−单磷酸;下调代谢物差异倍数从高到低的是顺−10−十七碳烯酸、高圣草酚、对香豆酰胆碱、橙皮素 5−O−葡萄糖苷、芥子碱、芹菜素−7−O−(6−O−丙二酰基葡萄糖苷)、茉莉酸、儿茶素−(7,8−bc)−4β−(3,4−二羟基苯基)−二氢−2−(3h)−吡喃酮、苜蓿素−7−O−二葡萄糖醛酸苷、2−脱氧核糖−5−磷酸。其中上调的代谢物中,2,6−二氨基庚二酸属于氨基酸及其衍生物,参与赖氨酸生物合成通路;下调的代谢物中,芥子碱属于生物碱,参与苯丙烷生物合成代谢通路。
胁迫5 d 与胁迫10 d 时相比,上调代谢物差异倍数从高到低的是龙胆酸、咖啡碱、2,6−二氨基庚二酸、尿苷 5'−二磷酸、根皮苷、5−甲基胞嘧啶、苯甲脒、15−羰基二十碳四烯酸、2,3−二羟基丙基−9,12−十八碳二酸酯−己糖−己糖、矢车菊素3−O−(3'',6''−二丙二酰葡萄糖苷);下调代谢物差异倍数按从高到低的是茉莉酸、高圣草酚、橙皮素5−O−葡萄糖苷、天竺葵素 3−O−葡萄糖苷、奎宁、芥子碱、高精氨酸、儿茶素−(7,8−bc)−4β−(3,4−二羟基苯基)−二氢−2−(3h)−吡喃酮、4−(氨甲基)−5−(羟甲基)−2−甲基吡啶−3−醇和茉莉酸−异亮氨酸。其中上调的代谢物中,矢车菊素 3−O−(3'',6''−二丙二酰葡萄糖苷)属于黄酮类,参与花青素生物合成通路;下调的代谢物中,茉莉酸属于有机酸,参与α−亚麻酸代谢和植物激素信号转导通路,橙皮素 5−O−葡萄糖苷、天竺葵素 3−O−葡萄糖苷和高圣草酚属于黄酮类,4−(氨甲基)−5−(羟甲基)−2−甲基吡啶−3−醇参与维生素 B6 代谢代谢,茉莉酸−异亮氨酸属于有机酸,参与植物激素信号转导途径。
所有出现在胁迫过程中的上调和下调排名前10 位的代谢物见表2~3。由表2~3 可知,上调的代谢物中包括了3 种黄酮、3 种脂质、2 种酚酸、2 种有机酸和1 种木质素和香豆素等;下调的代谢物中,包括了3 种生物碱、7 种黄酮、2 种脂质、2 种有机酸和1 种酚酸等。

表2 叶片不同处理间上调差异倍数排名前10 位的代谢物性质分析Table 2 Analysis of the top 10 metabolites in different leaf treatments by up-regulated fold difference

表3 叶片不同处理间下调差异倍数排名前10 位的代谢物性质分析Table 3 Analysis of the top 10 metabolites in different leaf treatments by down-regulated fold difference
3 结论与讨论
植物中含有众多代谢物,分为初级代谢物和次级代谢物。其中初级代谢物主要参与植物的基本生物合成过程,如三羧酸循环 (TCA)、糖酵解、脂肪酸的生物合成;还参与大分子的产生,如碳水化合物、脂质和蛋白质。植物生存不需要次级代谢物,如黄酮类化合物、抗氧化分子,主要是参与植物应对不良环境条件[15−16]。
香根草叶片在扑草净胁迫下测定出多种差异代谢物,主要包括氨基酸、有机酸、生物碱、脂质和黄酮等。其中氨基酸类物质在胁迫过程中呈差异变化的有:丙氨酸、蛋氨酸、高精氨酸等几种氨基酸在胁迫5 d 时呈上调趋势,鸟氨酸、组氨酸、天冬酰胺、异亮氨酸、焦谷氨酸等呈下调趋势;胁迫至10 d 时,天冬氨酸呈上调趋势,而苯丙氨酸、色氨酸、焦谷氨酸、半胱氨酸、高精氨酸等呈下调趋势。氨基酸是一类受环境压力影响的代谢物,参与多种代谢途径并可以合成许多次级代谢物,这些次级代谢物可能参与防御、信号传导等过程。氨基酸上调可能是由于蛋白质降解为单个氨基酸,并转化为参与三羧酸循环循环中用以产生能量的分子[17];另一方面,氨基酸下调可能是由于其被用于蛋白质的合成,合成用于应对胁迫环境的蛋白质组,从而导致下调;此外,植物中的一些氨基酸,尤其是脯氨酸,被用作渗透保护剂(也称为相容渗透剂),以应对受大多数非生物胁迫影响的细胞渗透压的变化。这些分子能够作为溶质而不干扰酶的活性、pH 值或是分子电荷,从而达到平衡渗透压的作用,但不会产生任何严重的附带效应[18]。香根草叶片的氨基酸在胁迫过程中下调数量多于上调数量,可见叶片内的氨基酸在此过程中主要用于合成应对胁迫环境的蛋白质组;而上调的代谢物种类中胁迫5 d 时有机酸的数量有所上升,可能是为了应对胁迫时产生的离子毒害而起到缓冲作用[19−20]。在差异代谢物通路中精氨酸和脯氨酸代谢通路在胁迫5 d 时被富集的7 种代谢产物有差异表达,谷胱甘肽代谢通路的4 种代谢产物有差异表达,应该是叶片在初期应对胁迫的主要代谢方式。
丙酮酸在胁迫5 d 时上调了2 205.44 倍,丙酮酸参与植物体内三羧酸循环等多个代谢途径,还是植物呼吸作用的潜在调节因子,能够刺激交替氧化酶(AOX)活性的增高。研究显示,外源添加丙酮酸和琥珀酸能提高植物欧李(Cerasus humilis)的呼吸作用,从而提高对干旱胁迫的抗性[21]。在胁迫5 d 时,香根草有7 种生物碱类物质呈上调;至10 d 时,又有2 种生物碱呈上调。其中五羟色胺及其衍生物N−阿魏酰五羟色胺在胁迫5 d时显著升高,有研究显示,在盐胁迫逆境处理时,五羟色胺及其酰基化衍生物的含量均有明显提高。综合本研究结果,推测其在非生物胁迫的响应过程中起到一定作用[22]。腐胺则是在胁迫10 d与5 d 相比时呈上调趋势,腐胺在植物胁迫时对于酶的稳定很重要,并可在避免由活性氧 (ROS)进行的氧化过程中发挥作用[18]。
植物激素通过充当信号分子来调节生物过程,以及上调或下调特定基因的表达。水杨酸 (SA)、茉莉酸 (JA)和脱落酸 (ABA)是植物在胁迫下产生的3 种最重要的激素物质[23],其有助于植物生长发育,还在信号传导过程中有信号分子的作用,促进多种代谢过程。本研究中香根草在扑草净胁迫下,叶片中茉莉酸在胁迫10 d 时比5 d 下调了13.27 倍,究其原因,植物的防御是一个高能耗过程,防御的适时抑制和终止有利于植物的生长发育。有研究表明[24],茉莉酸信号通路中通过降低茉莉酸或茉莉酸−异亮氨酸的累计使茉莉酸信号通路受到抑制,防止植物因防御过度而影响生长发育,本研究中茉莉酸和茉莉酸-异亮氨酸在胁迫后期均被检测到呈大幅度的下调表达。
综合分析表明,香根草叶片在胁迫过程中,下调代谢物数量要高于上调代谢物数量,且在胁迫0~5 d 时差异代谢物数量要高于胁迫5~10 d;丙酮酸、茉莉酸、五羟色胺等物质推测在抗逆过程中起主要作用,其中茉莉酸可能在胁迫后期起到防御信号终止的作用;游离氨基酸下调的数量要高于上调的数量。从代谢通路上看,精氨酸和脯氨酸代谢和植物激素信号转导可能在叶片抵御扑草净胁迫中起主要作用。
