Nrf2在黄芩素抑制氧化应激诱导心肌细胞凋亡中的作用
2022-06-15徐由财丁文俊陈俊邦陈婷芳曾科峰张竞之
徐由财, 丁文俊, 陈 思, 陈俊邦, 陈婷芳, 曾科峰, 刘 彬,2*, 张竞之,2*
(1.广州医科大学附属第二医院中医科,广东 广州 510260;2.广州医科大学中西医结合研究所,广东 广州 510180)
心血管疾病是世界人口死亡的主要原因[1],在我国发病率也逐年上升,其中缺血性心脏病是最受关注的问题之一[2]。氧化应激引起的心肌细胞损伤是缺血性心脏病发生的重要环节,它可引起动脉粥样硬化、心肌缺血、缺血再灌注损伤等[3],因此缓解心肌细胞氧化损伤和凋亡是防治缺血性心脏病的重要策略。
核因子E2相关因子(nuclear factor E2 related factor, Nrf2)是细胞抗氧化应激的关键转录因子[4],在一般情况下,Kelch样环氧氯丙烷相关蛋白1(Kelch-like ECH-associated protein 1,Keap1)会通过泛素化降解来抑制Nrf2的转录活性;在氧化应激条件下,Keap1的特定半胱氨酸残基被修饰,失去了泛素化降解Nrf2的能力,使其在细胞内积累并转移入核,诱导血红素氧合酶1(Heme oxygenase 1,HO-1)、醌氧化还原酶1[NAD(P)H quinone dehydrogenase 1,NQO1]等下游靶基因的表达。Nrf2信号通路在保护心肌细胞方面起着重要作用[5],其过表达能使小鼠维持氧化还原稳态,抑制氧化应激及心室重构[6]。
黄芩素是中药黄芩中活性最高的黄酮类化合物之一,具有抗氧化、抗炎、抗衰老、抗肿瘤等功效[7-8],Zhao等[9]发现,该成分能调节Ca2+水平影响蛋白的激活和表达,改善心功能及抑制细胞凋亡,但其心肌保护作用和潜在机制尚未明确。因此,本研究考察黄芩素抗氧化损伤、抗凋亡的作用及Nrf2信号通路在其中的作用。
1 材料与方法
1.1 试剂与药物 黄芩素对照品(批号MUST-19101105)购自成都曼思特生物科技有限公司。二甲基亚砜(DMSO,批号RNBK1113)、叔丁基过氧化氢(TBHP,批号BCCB8820)购自美国Sigma公司;胎牛血清(FBS,批号2029091)购自以色列Biological Industrie公司;DMEM培养基(批号8120120)、PBS(批号8120122)、胰蛋白酶(批号2072818)均购自美国Gibco公司;Nrf2抗体(货号12721)、IgG二抗(货号7074)购自美国CST公司;HO-1(货号AF5393)、NQO1(货号DF6437)、Bcl-2(货号AF6139)、Bax(货号AF0120)、β-actin(货号AF7018)抗体购自美国Affinity公司;SOD试剂盒(批号20201015)、GSH-Px试剂盒(批号20201014)购自南京建成生物工程研究所;BCA蛋白定量试剂盒(批号UJ292597)购自美国Thermo公司;MTS试剂(批号0000453287)购自美国Promega公司;TUNEL凋亡试剂盒(批号BZ2006024)购自武汉塞维尔生物科技有限公司;ML385抑制剂(批号S879001)购自美国Selleck公司。
1.2 细胞培养 H9c2细胞购自中国科学院上海细胞库,用含10% FBS、青霉素 (100 U/mL)、链霉素(100 mg/mL)的DMEM培养基,在37 ℃、5% CO2、饱和湿度的细胞培养箱中培养,倒置显微镜下观察细胞生长状态,当呈80%~90%融合时以0.25%胰蛋白酶消化传代,隔日换液,每2~3 d传代1次。
1.3 分组 本研究采用TBHP构造H9c2细胞氧化损伤模型,探讨黄芩素抑制TBHP诱导的H9c2细胞凋亡及氧化损伤时,分为对照组、模型组和黄芩素2.5、5、10 μmol/L组;探讨黄芩素能否直接激活Nrf2通路时,分为对照组和黄芩素2.5、5、10 μmol/L组;探讨ML385能否抑制黄芩素激活Nrf2信号通路时,分为对照组、模型组、黄芩素组(10 μmol/L黄芩素)、ML385组、黄芩素+ML385组(10 μmol/L黄芩素+20 μmol/L ML385)。
1.4 MTS法检测细胞活力 调整细胞密度至5×104/mL,接种于96孔板中,每孔100 μL,置于培养箱中培养24 h后加入含不同浓度待测药物的培养基,每孔100 μL,每组设4个复孔,放入培养箱培养12 h后造模2 h,弃旧培养基,每孔加入100 μL含MTS的工作液(DMEM∶MTS=5∶1),37 ℃恒温箱内避光孵育2 h,采用酶标仪在490 nm波长处检测光密度(OD)值,并计算细胞存活率。
1.5 分子对接及分子动力学 采用Autodock Vina软件[10]进行分子对接,分析Keap1(PDB ID:6QMC)与黄芩素(PubChem CID: 5281605)之间的结合相互作用。在分子对接前,去除原始空间结构中的水和配体来制备Nrf2的分子对接模拟蛋白,采用YASARA软件[11]进行分子动力学模拟。
1.6 表面等离子共振(SPR) 使用PlexArray HT系统进行表面等离子共振筛选以确定黄芩素和Keap1的结合,将Nrf2蛋白打印在3D传感芯片上,并通过光穿越反应将其固定,为获得结合亲和力,至少需要设置3个不同浓度。通过在25 ℃下以2 μL/s的体积流量流经样品进行300 s缔合,然后在运行缓冲液中进行300 s的解离,再以3 μL/s的体积流量运行200 s的再生缓冲液,可获得典型的结合曲线。
1.7 试剂盒检测细胞SOD、GSH-Px活性 将H9c2细胞以每孔2×105个的密度接种于6孔板,每组设3个复孔,待细胞长至80%~90%时分别给予不同浓度的药物预处理12 h,TBHP处理2 h,收集细胞,超声裂解,离心后取上清液,按照相关检测试剂盒说明书检测细胞SOD、GSH-Px活性。
1.8 TUNEL法检测细胞凋亡率 按“1.7”项下方法收集细胞,根据TUNEL细胞凋亡检测试剂盒说明书操作,在荧光显微镜下观察,计算凋亡率。
1.9 Western blot检测相关蛋白表达 按“1.7”项下方法收集细胞,PBS漂洗后用适量RIPA裂解液裂解15 min,刮取蛋白,4 ℃、12 000 r/min离心10 min,取上清,BCA蛋白定量试剂盒检测蛋白浓度,加入上样缓冲液,在100 ℃下加热10 min,12% SDS-PAGE进行电泳,转膜,脱脂奶粉封闭2 h,加入一抗4 ℃孵育过夜,PBST洗涤3次,每次7 min,加入二抗37 ℃孵育1 h,PBST洗涤3次,每次7 min,ECL化学发光试剂盒显色,扫描膜,采用Image J软件对蛋白条带灰度值进行分析,以目的蛋白与β-actin比值作为待测目的蛋白相对表达量。
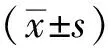
2 结果
2.1 黄芩素抑制TBHP诱导的H9c2细胞凋亡 如图1A所示,在0~600 μmol/L TBHP浓度范围内的细胞存活率随着浓度增加逐渐降低(P<0.01);与对照组比较,在TBHP浓度为200 μmol/L时,H9c2细胞存活率约为75%,故后续选取该浓度诱导H9c2细胞氧化损伤。如图1B所示,与对照组比较,模型组细胞存活率降低(P<0.01);与模型组比较,黄芩素预处理后细胞存活率提高并呈剂量依赖性(P<0.01)。如图1C~1D所示,与对照组比较,模型组H9c2细胞凋亡率升高(P<0.01);与模型组比较,黄芩素预处理后细胞凋亡率降低(P<0.01),并呈剂量依赖性。如图1E~1F所示,与模型组比较,黄芩素2.5 μmol/L组细胞Bcl-2/Bax比值表达无明显变化(P>0.05),而5、10 μmol/L组细胞Bcl-2/Bax比值表达升高,并呈剂量依赖性(P<0.01)。

注:与对照组比较,##P<0.01;与模型组比较,**P<0.01。图1 黄芩素抑制TBHP诱导的H9c2细胞凋亡Fig.1 Inhibitory effects of baicalein on TBHP-induced H9c2 cell apoptosis
2.2 黄芩素抑制TBHP诱导的细胞氧化应激 如图2所示,与对照组比较,模型组细胞上清液SOD、GSH-Px活性降低(P<0.05,P<0.01);与模型组比较,黄芩素2.5 μmol/L组细胞SOD、GSH-Px活性无明显变化(P>0.05),5、10 μmol/L组细胞SOD、GSH-Px活性升高,并呈剂量依赖性 (P<0.01)。

注:A~B分别为SOD、GSH-Px活性。与对照组比较,#P<0.05,##P<0.01;与模型组比较,**P<0.01。图2 黄芩素抑制TBHP诱导的氧化应激Fig.2 Inhibitory effects of baicalein on TBHP-induced oxidative stress
2.3 黄芩素与Keap1相互作用的分子模拟 本研究采用分子对接技术发现黄芩素能够与Keap1的Kelch位点结合,结合力为-9.0 kcal/mol,其三维结构如图3A所示,可知黄芩素与Keap1的氨基酸残基ASN-414、ASN-382、ARG-415、SER-508形成氢键结合。通过分子动力学模拟进一步研究了Keap1与黄芩素复合物的稳定性,在0、100 ns时的表面可视化模型如图3B所示,可知黄芩素能够稳定的呈现在Keap1结合位点的中心,直到分子动力学模拟结束。在开始的35 ns内,Keap1蛋白的均方根偏差(RSMD)从0.5 Å增加到1.5 Å,并围绕着1.25 Å波动(图3C,蓝色线),而黄芩素的RMSD稳定在0.5 Å波动(图3C,红色线),提示该成分与 Keap1结合是稳定的。
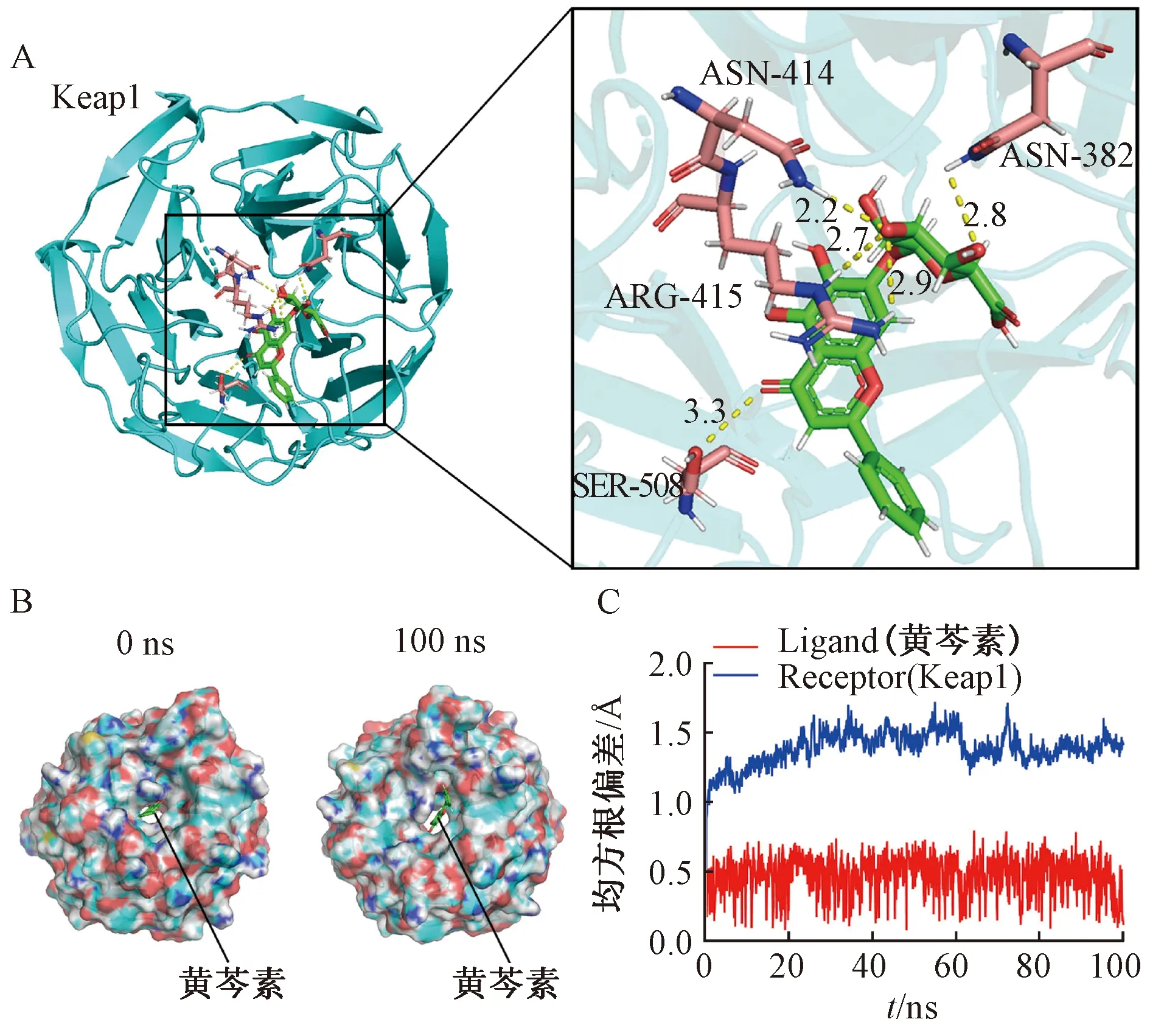
图3 黄芩素与Keap1相互作用的分子模拟结果Fig.3 Results for molecular simulation of the interaction between baicalein and Keap1
2.4 黄芩素与Keap1相互作用的表面等离子共振分析 如图4所示,黄芩素能与Keap1的Kelch位点直接结合,并且Keap1-黄芩素蛋白复合物能够稳定存在,并可以剂量依赖性地方式快速结合Keap1,平衡解离常数(KD)值为6.54×10-7mol/L,表明该成分对Keap1具有很强的亲和力,即与Keap1的Kelch位点结合较为稳定。
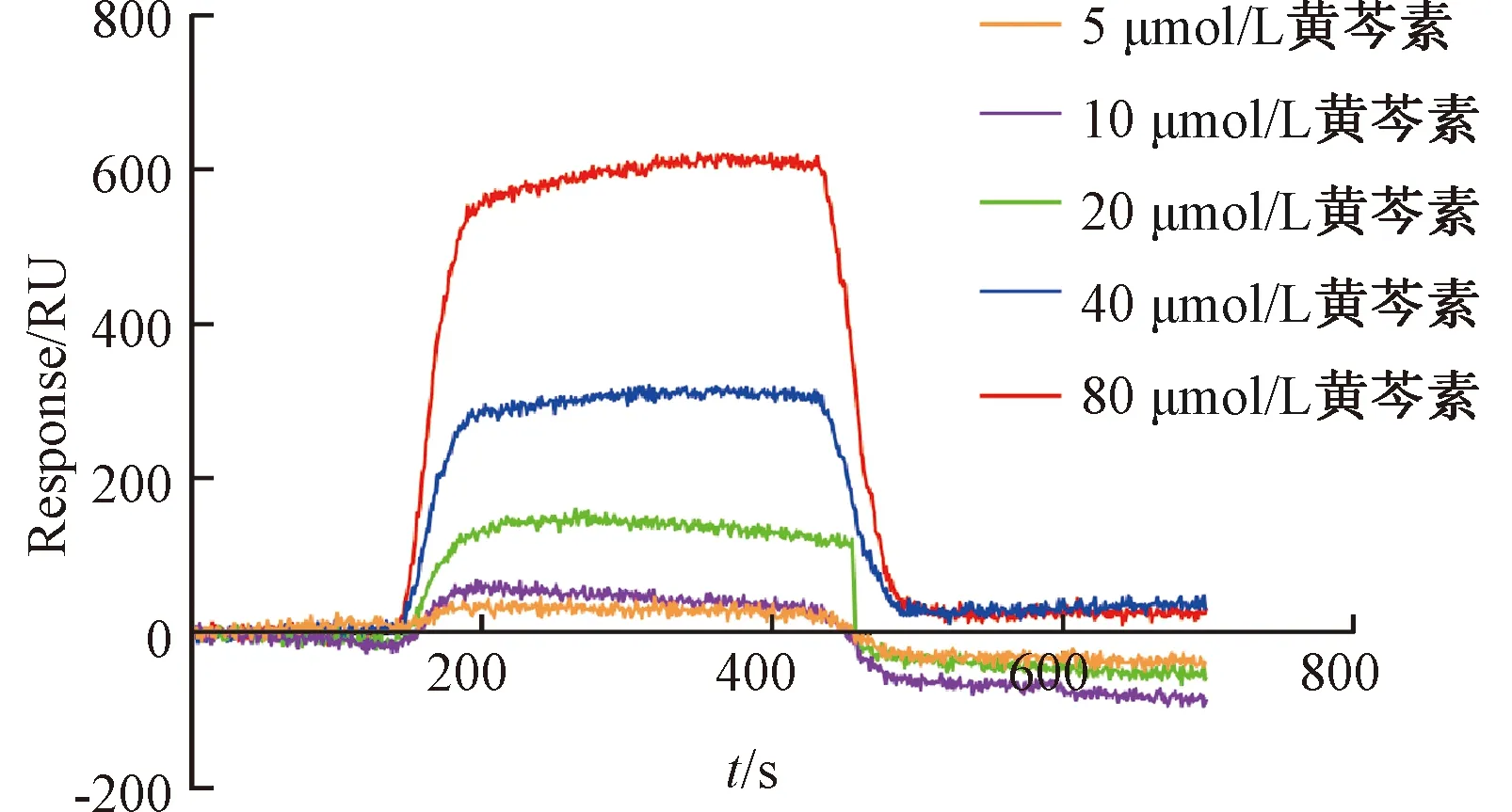
图4 黄芩素与Keap1相互作用的表面等离子共振结果Fig.4 Results for surface plasmon resonance of the interaction between baicalein and Keap1
2.5 黄芩素激活Nrf2信号通路 如图5所示,黄芩素能剂量依赖性地上调H9c2细胞内Nrf2的表达(P<0.01);与对照组比较,黄芩素2.5 μmol/L组细胞HO-1、NQO1蛋白表达无明显变化(P>0.05),5、10 μmol/L组细胞HO-1、NQO1蛋白表达升高,并呈剂量依赖性(P<0.01)。

注:A为Nrf2、HO-1、NQO1蛋白条带图,B~D分别为Nrf2、HO-1、NQO1蛋白表达统计图。与对照组比较,##P<0.01。图5 黄芩素激活Nrf2信号通路Fig.5 Activation of Nrf2 signal pathway by baicalein
2.6 ML385抑制黄芩素经Nrf2通路对心肌凋亡的保护作用 如图6所示,与黄芩素组比较,黄芩素+ML385组细胞存活率降低(P<0.01),细胞凋亡率、Bcl-2/Bax比值升高(P<0.01),提示ML385能够抑制黄芩素对心肌的保护作用。
2.7 ML385抑制黄芩素保护心肌免受氧化损伤的作用 如图7所示,与黄芩素组比较,黄芩素+ML385组细胞SOD、GSH-Px活性降低(P<0.01),Nrf2、HO-1、NQO1蛋白表达也降低(P<0.01),说明ML385能够抑制黄芩素保护H9c2细胞免受氧化损伤的作用。
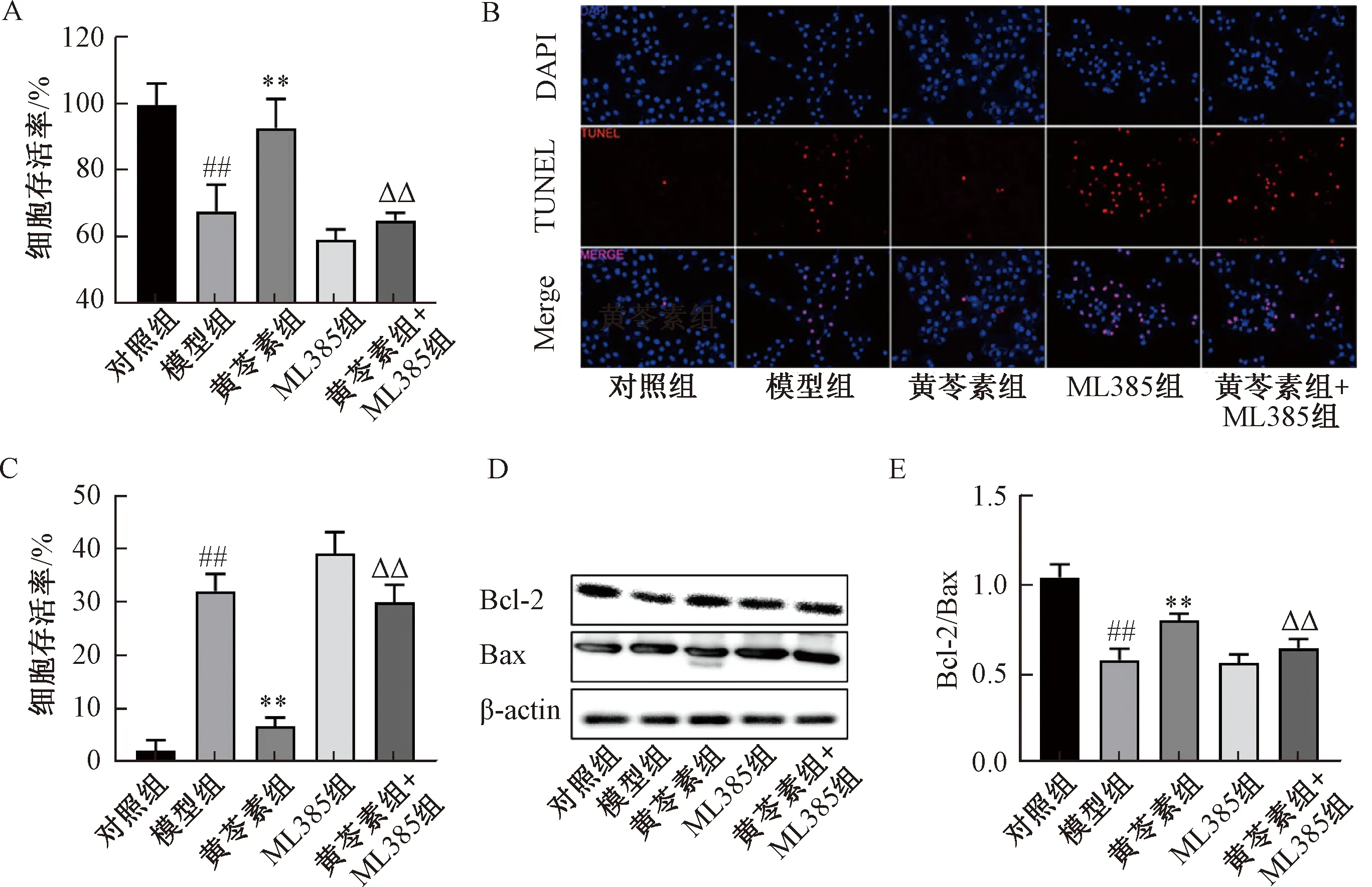
注:A为细胞存活率,B为细胞凋亡TUNEL染色,C为细胞凋亡率,D为Bcl-2、Bax蛋白条带图,E为Bcl-2/Bax的相对表达统计图。与对照组比较,##P<0.01;与模型组比较,**P<0.01;与黄芩素组比较,△△P<0.01。图6 ML385抑制黄芩素对心肌的保护作用Fig.6 Inhibitory effects of ML385 to baicalein’s protection on cardiomyocytes

注:A~B分别为SOD、GSH-Px活性,C为Nrf2、HO-1、NQO1蛋白条带图,D~F分别为Nrf2、HO-1、NQO1蛋白表达统计图。与对照组比较,#P<0.05,##P<0.01;与模型组比较,*P<0.05,**P<0.01;与黄芩素组比较,△△P<0.01。图7 ML385抑制黄芩素的抗氧化作用Fig.7 Inhibitory effects of ML385 on baicalein’s antioxidation
3 讨论
缺血性心脏病严重威胁人类生命健康,是人类死亡率及致残率最高的疾病之一。氧化应激是缺血性心脏病的发病机制之一,当机体受到某些刺激时,细胞内生成大量活性氧(ROS)与活性氮(RNS),一方面消耗抗氧化物质如SOD、GSH-Px等并生成有害物质如丙二醛(MDA)等,另一方面产生大量H2O2,并与金属离子反应生成高活性的羟自由基,引发细胞钙超载、蛋白质氧化、脂质过氧化,损害细胞膜完整性及功能,诱发细胞凋亡[12-13]。
黄芩素是中药黄芩的活性成分之一,在抗癌、抗炎、神经发生、心脏保护等方面均有较好的作用。王家伟等[14]认为黄芩素可激活JAK-STAT信号通路对大鼠心肌细胞缺血再灌注起到保护作用;Chang等[15]发现黄芩素能通过Akt-NOS-NO信号通路清除ROS的爆发,进而对缺血再灌注的鸡心肌细胞起保护作用。本研究发现黄芩素能提高TBHP诱导的H9c2细胞活力和SOD、GSH-Px活性,并减少细胞凋亡,具有良好的心肌保护作用。
在正常生理条件下,Nrf2基因表达的蛋白产物半衰期较短,缘于Keap1将Nrf2与E3泛素连接酶连接在一个复合物中,促进Nrf2蛋白的泛素化并通过26S蛋白酶体降解,这被认为是Nrf2负调控的典型机制[5]。Keap1是亲电子试剂和ROS的生物传感器,对细胞内Nrf2的调节起重要作用。Keap1主要具有BTB和 Kelch 两个保守结构域,其中BTB和泛素连接酶结合,Kelch与Nrf2结合[16]。由于缺乏Nrf2与Keap1结合位点的蛋白结构,目前Nrf2激动剂的药物研发都采用Keap1的Kelch位点虚拟筛选潜在的Nrf2激动剂。分子对接发现黄芩素能与Keap1的Kelch位点结合,结合力为-9.0 kcal/mol,并与Keap1的氨基酸残基形成氢键;分子动力模拟发现黄芩素与 Keap1结合是稳定的,SPR发现黄芩素与Keap1结合的平衡解离常数(KD)为6.54×10-7mol/L。这些结果提示黄芩素能与Keap1的Kelch位点结合,且这种结合能稳定存在。
本研究结果显示,黄芩素升高Nrf2、HO-1、NQO1蛋白表达,由此得出黄芩素可能是通过与Keap1的Kelch位点结合使Nrf2避免被泛素化降解,使其在细胞内迅速积累,转移入核并诱导下游靶基因的表达。Nrf2信号通路在心肌保护方面愈发受到重视,Jiang等[17]发现BRG1可上调Nrf2和HO-1表达来减轻急性心梗时心肌细胞的氧化应激;Hu等[18]发现益气活血方由Nrf2途径在心力衰竭时起到心脏保护作用,使H9c2细胞免受OGD/R诱导的凋亡;Cui等[19]发现虾青素经Nrf2信号通路对小鼠心脏氧化应激和赭曲霉毒素A暴露引起的线粒体凋亡起保护作用。本研究使用Nrf2抑制剂ML385后发现H9c2细胞活力和SOD、GSH-Px活性降低,细胞凋亡增加,说明ML385能抑制黄芩素对TBHP诱导心肌细胞凋亡的保护作用。
综上所述,黄芩素可能通过激活Nrf2信号通路减轻了TBHP诱导H9c2引起的氧化应激损伤和细胞凋亡,从而发挥其心肌保护作用。本研究为黄岑素在临床的运用提供了实验依据。
