不同大麦籽粒γ-氨基丁酸含量的差异及环境影响
2022-06-06张紫晋任益明粟永英钟志明
张紫晋,任益明,江 迪,杨 婷,粟永英,钟志明,陈 静
(1.中国科学院成都生物研究所, 成都 610041;2.中国科学院地理科学与资源研究所,北京 100101)
【研究意义】γ-氨基丁酸( γ-aminobutyric acid,GABA)是一种非蛋白质、功能性氨基酸,广泛分布于微生物和动植物体内的天然活性成分[1]。GABA仅存在于动物体内的神经组织,作为哺乳动物中枢神经系统中的抑制性神经递质,参与重要的脑循环生理活动,具有改善脑机能、增强记忆、抗焦虑等生理功能,同时具有抗心律失常、促进乙醇代谢、降低血压、预防肥胖、防止动脉硬化和改善糖尿病患者生理状况等功效[2-3]。随着年龄的增长和精神压力的增加,人体内GABA积累异常困难,需要从外界摄入一定量的GABA以满足机体的需要,富含GABA的谷物和豆类食品是理想的GABA膳食补充来源[4-6]。因此,开展谷物GABA的基因型和环境影响研究对于富含GABA的功能原料作物种植区划和品种培育具有重要意义。【前人研究进展】谷物γ-氨基丁酸的研究主要集中在水稻、玉米和大麦γ-氨基丁酸的富集培养及功能食品开发方面[7-9]。在冷激、热激、低氧、干旱等胁迫环境下,发芽籽粒GABA的含量显著增加,低温和低氧条件下谷物发芽是富集GABA的常用手段,研究人员因此开发了系列功能食品和饮料[10]。姚森等[11]发现水稻品种间GABA含量变异较大,平均GABA含量早稻品种高于晚稻,不同光温生态类型发芽糙米GABA的含量差异可能与水稻的品种特性和生育期内的光照、气温等环境条件有关。Makoto等[9]发现大麦发芽籽粒GABA含量显著高于水稻发芽籽粒,可成为γ-氨基丁酸的优质原料。赵大伟、曹彬等[12-13]分别对大麦品种类型、籽粒颜色与籽粒GABA含量差异进行了分析,发现裸大麦GABA含量高于皮大麦,籽粒GABA含量和籽粒颜色存在显著相关性,由于试验过程中的代表性供试材料数量不足,上述研究结果尚需进一步验证,而关于种植环境对大麦籽粒GABA含量影响的研究鲜有报道。【本研究切入点】大麦是重要的“药食同源”作物,尤其青藏高原裸大麦(亦称青稞)作为藏区人民的主粮,富含蛋白质、膳食纤维、维生素、花青素等营养健康成份,是高附加值食品或保健品开发的重要原料[14]。大多数品质性状,如籽粒蛋白质、淀粉组成、黄酮等功能成份,是受不同基因型,环境条件以及生长发育阶段影响的复杂性状[15],目前缺少大麦基因型和环境条件与籽粒GABA含量差异的系统研究,以及对富含GABA优良品种资源的挖掘利用。【拟解决的关键问题】本研究通过多点田间栽培试验,对大麦不同基因型、不同种植环境与籽粒γ-氨基丁酸含量的相关性进行评价,鉴定筛选优良种质资源,从而为GABA高含量的优质专用大麦种植规划和新品种培育提供科学依据和物质材料。
1 材料与方法
1.1 试验材料
本试验供试材料为358份栽培大麦品种,包含267份裸大麦和91份皮大麦材料,其中不同粒色裸大麦及份数见表1。2017—2019年将供试材料种植于四川什邡(104°09′E、31°10′N,海拔500 m,灌浆期气温21~28 ℃,降雨量281.7~327.5 mm)。2019—2020年,从什邡收获的材料中选取46份浅粒色(黄/白色)裸大麦和58份深粒色裸大麦,继续种植于云南元谋(101°35′~102°06′E、25°23′~26°06′N,海拔1100 m,灌浆期气温20~30 ℃,降雨量214.6~227.8 mm)和西藏拉萨(91°06′E,29°36′N,海拔3650 m,灌浆期气温12~24 ℃,降雨量302.2~344.7 mm),进一步了解种植环境对大麦籽粒GABA累积的影响。3个环境点种植,每一材料种植2行,行长1.5 m,行距0.25 m,株距3~5 cm。供试材料按随机区组排列,3次重复。采用当地常规水肥(底肥复合肥30~40 kg/667 m2,追施尿素5~15 kg/667 m2)及田间管理,于大麦完熟期收获种子。
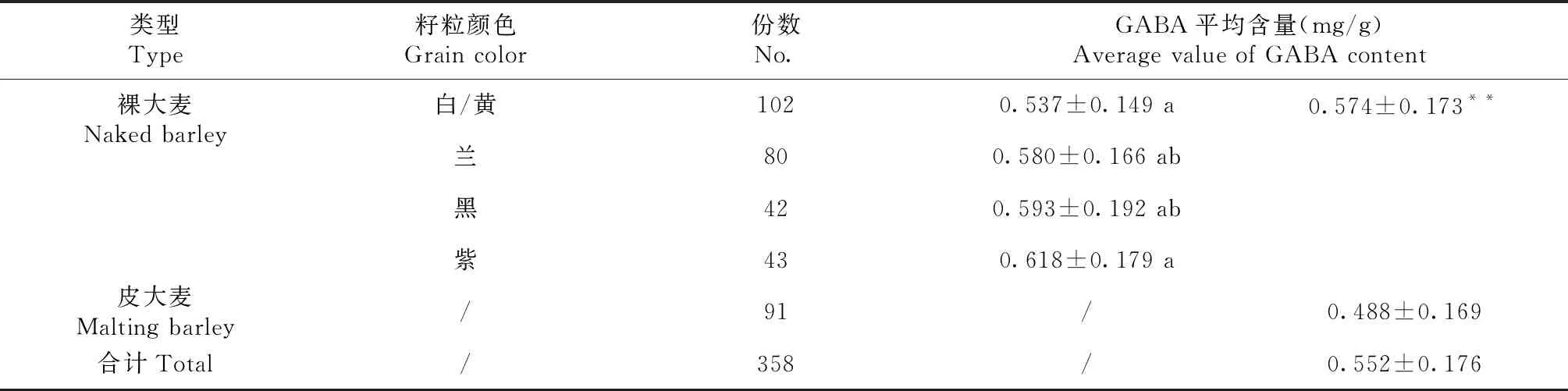
表1 供试大麦材料分组及GABA含量
1.2 试验方法
1.2.1 GABA标准曲线绘制 参考Tojiro等[16]的比色方法,稍作改动。称取10 mg GABA标准品,加入10 mL蒸馏水配制成1.0 mg/mL GABA标准溶液。用移液枪吸取0.00、0.05、0.10、0.15、0.20和0.25 mL标准溶液于10 mL的带盖试管内,并分别加入蒸馏水1.00、0.95、0.90、0.85、0.80、0.75 mL使每管溶液最终体积达到1 mL,得到不同浓度梯度的GABA标准溶液。向GABA标准溶液中依次加入0.2 mol/L硼酸盐缓冲液(pH 9.0) 0.6 mL、5%苯酚溶液2 mL、7%次氯酸钠溶液1 mL,摇匀后沸水浴加热5~10 min,直到颜色变成蓝绿色,放入冰浴冷却至室温,最后加入60%的乙醇2 mL摇匀静置。在波长645 nm处测定溶液的吸光度。以吸光度为横坐标,GABA含量为纵坐标,绘制标准曲线。
1.2.2 γ-氨基丁酸的提取及含量测定 将样品籽粒于70 ℃烘干后粉碎,过400目筛备用。准确称取150 mg面粉置于2 mL离心管中,加入1.5 mL蒸馏水于MIX-25P迷你混合仪上震荡浸提4 h。12 000 r/min离心5 min,取1 mL上清液于10 mL的带塞试管内,按照1.2.1方法显色反应后在645 nm下测吸光值,通过标准曲线计算出样品GABA的含量。
1.3 数据处理分析
采用Excel 2007进行试验数据整理和图表制作,利用SPSS 11.5软件进行统计分析, LSD方法对各样本平均数进行差异显著性检验。
2 结果与分析
2.1 标准曲线
将不同浓度梯度的GABA标准溶液,经显色反应后于室温下645 nm波长处测定吸光度(图1)。结果发现,GABA含量(y)与645 nm处吸光度(x)具有以下线性关系:y=0.0965x-0.0001,其R2=0.9989,表明该方法准确可靠,可用于供试材料的GABA含量测定。

图1 GABA标准曲线
2.2 大麦品种间籽粒GABA含量的差异
什邡种植点358份供试材料的籽粒GABA含量变异幅度较大(0.062~1.038 mg/g),平均值为0.552±0.176,大部分材料GABA含量位于0.45~0.66 mg/g,占总量的61%,表现为数量性状的正态分布特点(图2)。如表1所示,裸大麦平均GABA含量为(0.574±0.173)mg/g(变异幅度0.215~1.038 mg/g),皮大麦平均GABA含量为(0.488±0.169)mg/g(变异幅度0.062~1.014 mg/g),前者GABA含量极显著高于后者(P<0.01)。根据籽粒颜色,将裸大麦材料划分为黄/白粒、兰粒、紫粒和黑粒4组,每组对应的平均GABA含量高低顺序为紫粒>黑粒>兰粒>黄/白粒,其值分别为(0.618±0.179)、(0.593±0.192)、(0.580±0.166)、(0.537±0.149)mg/g,表明籽粒颜色越深GABA含量越高,其中紫粒GABA含量极显著高于黄/白粒,而兰粒与黑粒差异不显著。
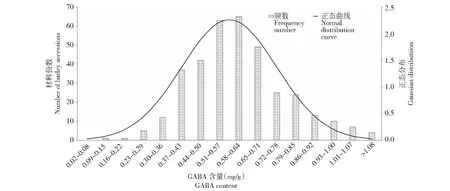
图2 不同大麦品种(系)GABA含量分布(什邡)
2.3 不同种植环境下裸大麦GABA含量的变化特征
从供试材料中选取46份浅粒色(黄/白色)裸大麦和58份深粒色裸大麦,在地理位置相近但海拔差距极大的四川什邡、云南元谋和西藏拉萨3个地点继续种植。从图3可以看出,同一批供试材料在不同种植地点的GABA含量集中分布范围不一样,在四川、云南和西藏种植点大多数材料的GABA含量范围分别为0.4~0.69、0.7~0.89和0.8~0.99 mg/g,而且GABA高含量(≥1.0 mg/g)的材料在四川仅有2份,而云南有27份,西藏达到32份,表明种植地点海拔高度对大麦籽粒GABA含量有正向促进作用。如图4所示,裸大麦浅粒色组平均GABA含量分别为(0.496±0.133)、(0.765±0.191)和(0.907±0.259)mg/g,深粒色组平均GABA含量分别为(0.603±0.179)、(0.864±0.231)和(0.917±0.312)mg/g。统计学分析表明,供试材料3个种植点的平均GABA含量随海拔高度增加明显提高,含量高低顺序为:西藏拉萨>云南元谋>四川什邡。其中浅粒色组GABA含量变化与总体趋势一致,表现为随环境海拔升高含量增加,且差异均达显著水平;深粒色组GABA含量也表现出随海拔升高含量增加的变化趋势,而云南与西藏2个环境点之间差异未达到显著水平。进一步分析发现,在四川和云南2个种植地点,深粒色组平均GABA含量均显著高于浅粒色组,分别高出21.73%和12.91%,而在西藏种植点2个粒色组之间没有差异,表明在相对低海拔的区域,裸大麦GABA含量受籽粒颜色的影响较大,而在高海拔地区受海拔高度的影响更显著。上述结果表明,大麦(青稞)籽粒GABA含量是受遗传控制和环境强烈影响的复杂数量性状。值得一提的是,通过2年的重复试验,我们从平均GABA含量>0.8 mg/g的材料中筛选到4份材料表现出较好的环境稳定性,其GABA含量在不同环境点之间差异不大(表2)。

图3 3个环境种植点裸大麦GABA含量分布
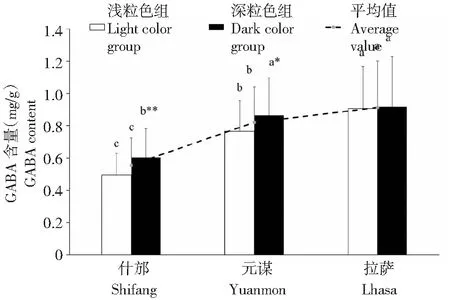
图4 3个环境种植点裸大麦平均GABA含量
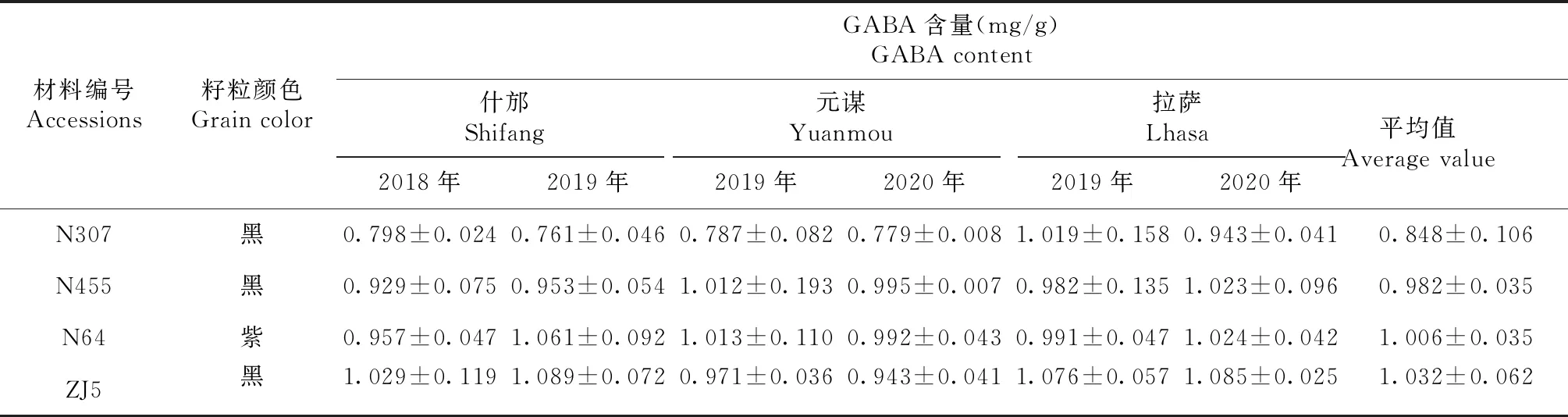
表2 环境稳定GABA含量较高的大麦品种
3 讨 论
γ-氨基丁酸广泛存在于各种植物及其不同组织中,担任着代谢物质和信号分子的双重角色,参与了植物的pH调节、能源物质调节、C/N平衡调节以及防御系统调节,在植物生长发育过程中起着重要的生物学保护功能[17-19]。相关研究表明,低温、低氧等环境胁迫能显著增加发芽谷物GABA含量[20],大麦和小麦耐低温冷害胁迫能力与GABA含量密切相关,耐寒品种GABA含量显著高于不耐寒品种[21]。施用外源GABA能提高植物抗氧化防护酶活性,有效缓解酸性土壤中铝离子对根系带来的氧化伤害,通过增加脯氨酸和酚类等物质的含量,以及提高植物抗氧化系统能力从而增强其盐胁迫耐性[22-24]。与前人研究结果一致[12-13],本研究供试大麦不同基因型间GABA含量变异较大,且裸大麦显著高于皮大麦,深色籽粒高于浅色籽粒。以谷氨酸或谷氨酸盐为底物的谷氨酸脱羧酶催化作用是GABA合成的主要途径,本研究证实大麦籽粒GABA含量与籽粒颜色密切相关,表明GABA合成途径可能与粒色相关的代谢途径存在交叉作用。进一步研究发现,总体上海拔高度对两组粒色裸大麦的GABA含量有明显的正向促进作用。在海拔相对较低的四川、云南种植点,深粒色组平均GABA含量显著高于浅粒色组,表明籽粒颜色对GABA含量的影响较大。随着环境海拔升高,在西藏种植点深粒色组GABA含量增幅较小,而浅粒色组增幅显著且几乎等同于深粒色组,表明在高海拔环境下海拔高度是影响大麦GABA含量的主要环境因子,而籽粒颜色对GABA含量的影响减弱甚至消失。因此,高原农区独特的自然环境条件有利于大麦籽粒GABA的累积,这为优质专用大麦种植区划和产业发展提供了科学依据。

籽粒GABA含量不仅受种子发育阶段温、光、水等环境条件的影响,还受到复杂遗传系统的控制。生物体内GABA的合成途径主要来自GABA支路[26],即谷氨酸经限速酶谷氨酸脱羧酶(Glutamate decarboxylase, GAD)催化脱去α位上的羧基形成GABA。水稻不同品种发芽糙米GABA生成量与GAD的酶活性呈极显著正相关[11],本研究发现大麦品种间GABA含量还与GAD基因表达水平相关(未发表数据)。本试验鉴定筛选到的GABA含量高且受环境影响小的大麦材料,为后续开展高GABA含量的大麦新品种培育及分子机制研究提供了重要的资源材料。
4 结 论
采用比色方法,对在什邡、元谋和拉萨不同地点种植的358份大麦材料籽粒γ-氨基丁酸含量进行了检测与分析。研究发现,大麦不同基因型之间GABA含量差异较大,裸大麦GABA含量显著高于皮大麦。在相对低海拔的环境下籽粒颜色与GABA含量密切相关,海拔高度对GABA含量有正向促进作用,是高海拔环境下影响籽粒GABA含量的主要环境因子。大麦(青稞)籽粒GABA含量是受遗传控制和环境强烈影响的复杂数量性状,同时鉴定筛选到4份环境表现稳定的GABA高含量资源材料。研究结果为优质专用大麦品种培育和种植区划提供理论依据和物质材料。
