尖峰岭热带山地雨林豆科树木对邻体树种的影响
2022-05-24邓方立林明献李意德
邓方立,许 涵,陈 洁,林明献,李意德
(1. 中国林业科学研究院热带林业研究所,广东 广州 510520;2. 南京林业大学,江苏 南京 210037;3. 中国林业科学研究院热带林业研究所试验站,海南 乐东 572542)
除人类活动之外,大气中的氮气进入生物圈主要有两个过程:闪电固氮和生物固氮(BNF)[1]。在原始的生物圈中,通常认为生物固氮作用比闪电固氮作用大一个数量级,所以,生物固氮被认为是固定氮的主要来源[2]。生物固氮除了细菌和真菌的固氮作用外[3],主要来源于豆科植物的生物固氮,并被认为是土壤氮的主要来源[4]。豆科植物固定大气氮的能力主要来自于与根瘤菌、土壤中的细菌的共生。豆科植物通过光合作用向根瘤菌提供能量和碳(C),根瘤菌主要以铵的形式向豆科植物提供氮[5]。豆科植物被认为是初级生产力、固碳、氮累积和矿化的关键驱动力[6-7],可以提高植物生产力[8-9]、氮的利用率[10]和土壤有机质的含量[11]。热带森林中,尽管土壤中矿物氮相较于温带森林很高[12],生物固氮率仍然保持高固氮水平,达到15~36 kg·a−1·hm−2。这种高生物固氮率的保持不是为了获取氮本身,而可能是为了获取磷或其它限制性养分酶的产生需要很高的氮输入[13]。
豆科植物的固氮作用还会对邻体植物的生长产生影响。在农作物的研究中,通过间作,大豆可以通过根系分泌物输出,直接向邻体植物提供氮,并通过菌根活动进一步促进磷的移动[14-15]。但是,在森林群落中豆科植物对非固氮树种生长和对整个森林的初级生产力的影响是不明确的。一方面,豆科固氮植物可以通过向非固氮树木提供可利用的氮,并提高森林的生产力。Temperton 等[16]认为,固氮植物与非固氮植物之间的相互促进作用的主要驱动力是减少氮竞争的氮节约,而氮转移则起次要作用。豆科植物转移到邻体植物的氮量可能占非豆类植物全氮的8%~39%[17],这取决于群落的年龄[18]。豆科植物还会通过共生菌根直接利用大气氮气,大大增加生态系统中氮的输入,形成群落中固氮物种和非固氮树种之间的互补[19]。另一方面,豆科固氮树种可能会抑制邻体非固氮树种的生长。因为,豆科树木通常具有更快的生长速度,有明显的竞争优势,使邻体物种多样性降低,长期影响下也可能降低邻体树木存活率。同时,有些豆科植物也会通过化感作用来抑制其他物种的生长,并抑制发芽和幼苗的形成[20]。另外,豆科固氮树种自身的迅速生长还可能对其他非固氮树种产生中性影响,从而导致森林总生物量的增加[21]。
因此,豆科树木对森林和其邻体的作用具有不确定性。为了进一步明确豆科树木在热带山地雨林中的角色和对邻体的作用,本文以尖峰岭60 hm2大样地内7 种共9 232 株豆科树木为研究对象,分析豆科树木0~16 m 邻体距离范围内、2 次调查间邻体非豆科树木的物种多度和丰富度的变化及邻体存活率,揭示了豆科树木对邻体树种多度、丰富度和存活率的影响,为探索豆科树木与邻体树木的共存机制提供依据。
1 研究区概况
尖峰岭位于海南省西南部,位于乐东县与东方市交界处(18°20′~18°57′ N,108°41′~109°12′ E),面积约640 km2。林区内的热带雨林是我国现有面积较大、保存较完整的热带原始森林之一。尖峰岭地区属低纬度热带岛屿季风气候区,年均降水量2 449 mm,全年蒸散量占35.7%~51.4%[22]。具有明显的干湿两季,年均气温24.5 ℃。尖峰岭地区是我国生物多样性较高的地区之一,野生维管植物和栽培植物共2 849 种,其中,野生植物2 287 种(包括变种、亚种和变型),隶属于224 科,1 250 属[23],热带山地雨林组成复杂,大多数种群的生态位宽度值较小[24]。另外,该地土壤为砖黄壤-黄壤,土壤各物理性质空间异质性明显[25]。
2 研究方法
2.1 样地设置及调查
2012 年在海南尖峰岭热带山地雨林区的五分区原始林内,参照美国史密森热带研究所热带森林研究中心(CTFS)的调查技术规范[26],建立一个东西长1 000 m,南北宽600 m,面积为60 hm2的样地(简称尖峰岭大样地),整个样地被分为1 500个20 m × 20 m 的样方。样地内海拔在866.3~1 016.7 m 间变化。
尖峰岭大样地首次植被调查开始于2010 年年底,2012 年完成,调查结果显示,尖峰大样地中共有胸径>1 cm 的活体植株439 676 株。第1 次复查于2018 年年底完成,共记录到胸径>1 cm的植株441 551 株(表1)。尖峰岭60 hm2大样地中共有7 种豆科树木:薄叶猴耳环、猴耳环、亮叶猴耳环,肥荚红豆、木荚红豆、软荚红豆、长脐红豆(表2)。这7 种豆科树木均是具有生物固氮能力的树种。豆科树木的固氮能力可以通过豆科植物与其邻体植物叶片中的δ15N 含量相比较计算得出[27]。

表1 尖峰岭大样地内多度前10 的木本植物Table 1 Top 10 woody plants with the highest abundance in JFL 60 hm2plot

表2 尖峰岭大样地豆科树木基本情况Table 2 Basic information of legume trees in JFL 60hm2plot
2.2 数据处理
2.2.1 邻体多度、丰富度和存活率计算方法
多度(N):N=邻体树木个体数
丰富度(S):S=邻体树木物种数
存活率(SR):SR=N2/N1
式中:N1为初次调查时某一范围内的个体数,N2为复查时该范围内依然存活的个体数
为了评估特定豆科物种的存在是否对邻体树木的多度、丰富度和存活率有显著影响,本文采用基于个体与物种的区域关系,简称ISAR(individual species-area relationship)[28],进行点标记模式统计分析。ISAR被定义为距焦点物种i的距离r以内的树木多度、丰富度或存活率的变化:

式中:Sij(r)是在距离r范围内的物种i的j个体周围的物种多度、丰富度或存活率,Ni是物种i的单个个体的数量,在这里指尖峰岭大样地中的7 种豆科树木。
为了检验豆科树木邻体相对多度、丰富度或存活率是否显著高于或低于非豆科树木,计算邻体非豆科树木的ISAR与豆科树木做比值(RISAR):

式中:Hij(r)为邻体非豆科树木的邻体多度、丰富度或存活率。这里邻体非豆科树木的选取标准是:与豆科树木生长在同一相似生境范围(距离豆科树木60 m 内)、且处于同一径级(径级为5 cm)的非豆科树木,且r=2, 4 , ….,16 m。理论上,距离中心豆科树木距离越近,生境越相似,本文选择距离豆科树木60 m 的原因是,在尽量保证生境相似前提下,保证统计有相当数量的非豆科树木可以选出作为对比。
如果RISARi(i)>1,表示豆科树种与同一生境下、同一径级内的非豆科树木相比,对邻体多度、存活率或丰富度有促进作用。如果RISARi(i)<1,表示豆科树种与同一生境下、同一径级内的非豆科树木相比,对邻体的物种多度、存活率或丰富度有抑制作用。
2.2.2 邻体多度、丰富度和存活率的影响因素分析 离豆科树种的距离越近,邻体影响越明显。2 m 和4 m 是豆科树木对邻体影响最有可能起作用的距离,所以,本文单独选出2 个最小尺度进行分析。为了进一步检验邻体多度、丰富度和存活率之间的关系及可能的影响因素,利用尖峰岭大样地内7 种豆科树种的叶片氮含量、固氮能力[27]、2 次调查中豆科树木2 m 和4 m 范围内的邻体多度、丰富度和存活率进行一元线性回归相关分析。
所有数据分析和绘图均通过R 4. 0. 2 (R Development Core Team, 2020)进行。
3 结果与分析
3.1 随邻体距离增加豆科树木的邻体相对多度变化
在邻体距离0~16 m 内,2012 年软荚红豆和亮叶猴耳环的邻体多度均大于非豆科树种的邻体多度,长脐红豆、肥荚红豆、薄叶猴耳环和木荚红豆的邻体多度均小于非豆科树种的。在邻体距离0~8 m 内,猴耳环的邻体多度小于非豆科树种的;在邻体距离8~16 m 内,猴耳环的邻体多度大于非豆科树种的(图1)。
在邻体距离0~16 m 内,2018 年只有软荚红豆多度大于非豆科树种的,亮叶猴耳环、荚红豆、薄叶猴耳环和木荚红豆的邻体多度均小于非豆科物种的。在邻体距离0~6 m 内,猴耳环的邻体多度小于非豆科树种的;在邻体8~16 m 内,猴耳环的邻体多度大于非豆科树种的(图1)。

图1 尖峰岭60 hm2大样地豆科树木的邻体相对多度Fig. 1 Relative neighboring abundance of leguminous trees in JFL 60 hm2plot
3.2 随邻体距离增加豆科树木的邻体相对丰富度变化
在邻体距离0~16 m 内,2012 年软荚红豆、亮叶猴耳环和猴耳环的邻体相对丰富度均大于非豆科树种的,长脐红豆、肥荚红豆、薄叶猴耳环和木荚红豆的邻体相对丰富度小于非豆科树种的。2018年结果与2012 年相似(图2)。
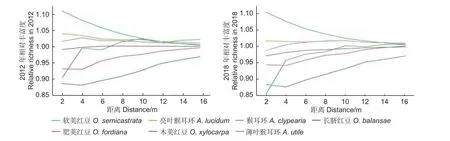
图2 尖峰岭60 hm2大样地豆科树木的邻体相对丰富度Fig. 2 Relative neighboring richness of leguminous trees in JFL 60 hm2plot
3.3 随邻体距离增加豆科树木的邻体相对存活率
在邻体距离0~16 m 内,软荚红豆和猴耳环的邻体相对存活率均大于非豆科树种的,亮叶猴耳环、木荚红豆、长脐红豆和薄叶猴耳环的邻体相对存活率均小于非豆科树种的。在邻体距离0~4 m内,肥荚红豆的邻体相对存活率大于非豆科树种的;在邻体距离6~16 m 内,肥荚红豆的邻体相对存活率小于非豆科树种的(图3)。

图3 尖峰岭60 hm2大样地豆科树木的邻体相对存活率Fig. 3 Relative neighboring survival rates of leguminous trees in JFL 60 hm2plot
3.4 相对多度和丰富度在邻体距离2 m 和4 m 的动态变化
2 m 和4 m 是豆科树木对邻体最可能的影响距离。在邻体距离2 m 和4 m 时,2012 年和2018年2 次调查,豆科树木邻体相对多度和丰富度无显著变化,7 种不同豆科树木邻体其他种树的多度和丰富度的影响结果保持不变(图4)。

图4 豆科树木邻体2 m 和4 m 的相对多度和丰富度变化Fig. 4 Changes of relative abundance and richness of leguminous tree in 2 m and 4 m radius from 2012 to 2018
3.5 豆科树木邻体相对多度、丰富度和存活率的影响因素分析
从表3 可看出:在尖峰岭60 hm2大样地中,豆科树木的邻体多度和丰富度有着显著的线性相关关系(p<0.05),但是邻体多度、丰富度与自身的叶片氮含量和固氮能力无显著的关系。存活率方面,只有邻体距离为4 m 范围的存活率与叶片氮含量有显著的线性相关关系(p=0.042 5)。

表3 7 种豆科树木的邻体多度、丰富度、存活率与叶片氮含量、植株固氮能力的相关关系Table 3 Relationship among neighbor abundance, richness, survival rates, leaf nitrogen content and nitrogen fixation abilities of seven leguminous trees
4 讨论
以上研究结果表明,豆科树木对邻体的影响与物种本身属性有关,软荚红豆和猴耳环对邻体存在正向的促进作用,薄叶猴耳环、木荚红豆、长脐红豆和肥荚红豆树木对邻体存在负向的抑制作用,亮叶猴耳环对邻体产生中性影响。在其他的研究中也有类似发现,如在草原和农田生态系统中,禾本科草本植物的相对产量和密度受豆科牧草混播比例和种类的影响[29-30]。在森林生态系统中,豆科树种对其他树种的影响不尽相同,如在次生林演替中,豆科树木的丰富度对生物量的恢复没有影响[21];在原始林,豆科树木对邻体多样性存在正或负的影响[31]。因此,豆科对邻体的影响会从多个方面进行影响,包括邻体的生物量、物种多样性、存活率等,并且影响效果存在差异。从2012 和2018的2 次调查的时序动态变化看,7 种豆科树木对邻体相对多度、丰富度的正负影响保持不变,相对稳定,只是影响幅度发生变化,即RISAR 发生变化。
本研究中,软荚红豆的邻体多度、丰富度和存活率均大于非豆科树种的,这表明在尖峰岭大样地中软荚红豆可能是贡献者,能对邻体树木产生正向的促进作用。结合尖峰岭大样地内豆科树种的叶片N 含量和固氮能力分析[27],软荚红豆具有最低的叶片N 含量和中等的固氮能力,软荚红豆可能是通过向土壤或是直接向邻体树木提供N 的方式来促进邻体树木的生长。薄叶猴耳环和木荚红豆的邻体多度、丰富度和存活率在数据上低于参照的非豆科树种的,表现出对邻体的抑制作用。这可能是薄叶猴耳环和木荚红豆的种群数量较小,且集中于小径级。稀有种通常占据边缘生态位,来避免与优势树种的竞争[32]。所以,薄叶猴耳环和木荚红豆邻体较低的相对多度、相对丰富度和相对存活率可能并不来自于树种本身,而是因为它们邻体范围内的环境压力更大。
已有研究发现,尖峰岭大样地中豆科树木的生物固氮能力与其所处土壤的速效氮含量紧密相关,在氮富集的环境中,豆科树种会表现出更高的生物固氮能力和邻体多样性[27];同时,在全球其它10 个样地的研究结果也显示,在土壤氮含量高的环境中,豆科树木的邻体有更高的多样性,而在低土壤氮环境下,豆科树木的邻体多样性更低[31]。也就是说尖峰岭大样地中,豆科树木在高氮环境中会有更高的固氮能力,在低氮环境下固氮能力更低。这就说明了豆科树木对邻体存在正或负影响的可能原因是:在高土壤氮条件下,土壤中细菌和真菌的多样性会增加[33-34],这些细菌和真菌会加速豆科富含氮掉落物的矿化分解[35-36],这些氮元素会进入土壤,被邻体其它植物利用,而导致相对高氮条件下豆科树木邻体更高的多样性和多度。此条件下,氮元素的循环快,豆科树种起到一种氮元素重新分配的作用。相反,在低土壤氮条件下,豆科树种降低自身的固氮效率,形成一种低氮耐受的生活方式,豆科树木在氮限制的环境中就会有强烈的竞争优势。此条件下,氮元素的循环慢,豆科树木同自身的竞争优势而导致邻体的多度和丰富度降低。总而言之,豆科树种对邻体的影响不仅取决于豆科物种的种类,而且这种竞争能力会随土壤养分和光照等资源的可用性,尤其是土壤氮的变化而变化[37]。
5 结论
总的来说,从现有研究结果看,豆科树木在热带森林中的角色是复杂多变的。这与在其它地区研究结果相似,也说明了豆科植物在不同的生态系统中作用是不同的。从尖峰岭大样地分析结果看,软荚红豆对邻体树木有明显的促进作用,未来可以选做造林混交树种使用。基于现有的数据和不同地区的研究结果,热带森林中豆科树木与邻体之间可能存在根际微生物的相互作用,同时受土壤养分的影响。接下来需要对豆科树木对邻体植物生物量和生长速率的影响进行分析,并且开展相应的控制试验,并探索其与根际微生物共存相关的生长表现,以系统的揭示豆科树木对邻体树木的影响及其内在机制。
