干旱胁迫对春小麦旗叶生理特征及其根系抗旱基因表达特征的影响
2022-05-24史功赋魏淑丽程玉臣张向前王建国赵小庆路战远
方 静,史功赋,魏淑丽,程玉臣,张向前,王建国,安 玉,赵小庆,路战远
(1. 内蒙古大学生命科学学院,内蒙古 呼和浩特 010020;2. 内蒙古自治区农牧业科学院,内蒙古 呼和浩特 010031;3. 内蒙古自治区退化农田生态修复与污染治理重点实验室,内蒙古 呼和浩特 010031)
干旱是影响作物生长发育最主要的自然灾害[1],其导致作物减产量已经超过其他因素造成的减产量总和[2-5]。小麦(TriticumaestivumL.)是三大主粮之一,我国小麦的种植面积及产量仅次于玉米、水稻[6-7]。呼伦贝尔市是内蒙古自治区重要的商品粮输出基地[8],春小麦是该地区主要种植作物之一,分布范围较广,种植历史较长。其中,在呼伦贝尔市陈巴尔虎旗、新巴尔虎左旗、牙克石市等大兴安岭西麓旱作区春小麦种植面积最大,约占全市春小麦种植面积的82%[8]。因此,对干旱胁迫下春小麦生理特征及其根系基因表达情况的变化特征进行研究,以期为进一步量化春小麦受旱程度和旱区作物抗旱性研究提供理论依据和技术路径。
光合作用对作物生长发育和生产力起到决定性作用[9-10]。干旱胁迫会导致春小麦旗叶光合及生理生化指标发生改变。张士昌等[11]研究发现,干旱胁迫下抗旱小麦品种产量高于水敏感品种,且抗旱小麦品种旗叶光合速率、蒸腾速率和气孔导度下降幅度均低于水敏感品种。郭程瑾等[12]研究不同小麦品种的光合特性发现,干旱胁迫对抗旱性强的品种光合影响要比抗旱性弱的品种低。也有研究表明,土壤含水量是气孔开启程度的决定因素,土壤水分受限制时小麦叶片气孔导度降低[13]。植株受到干旱胁迫时其生理特征也会发生显著变化。前人研究发现植物受到干旱胁迫时,其细胞内清除活性氧的酶促体系(SOD、POD等)、细胞渗透调节物质(Pro)含量均有升高趋势,能够清除植物机体内活性氧有害物质并减轻叶片组织失水,进而提高作物的抗旱性能[11,14]。
植物生长发育和对逆境的诱导反应涉及到复杂的基因表达调控网络[15]。研究干旱胁迫下基因表达变化规律是揭示干旱胁迫分子机理的重要手段[15-16]。不同基因型春小麦耐旱程度存在着极大的差异性[17-19],且不同春小麦品种对干旱胁迫响应机制和调控途径也存在较大差异。根系作为植物的水分吸收器官,其基因表达变化可直观反映出作物抗旱性的强弱[18-19]。周琪等[20]研究表明,干旱处理下小麦抽穗期根、茎、叶和穗中木葡聚糖内转糖苷酶基因(TaXTH-7A)均有表达,并在根中优势表达。李素等[21]通过在小麦两叶一心期连续进行4 d PEG胁迫处理条件下,进一步筛选小麦抗旱基因,TaBADHb基因呈上调趋势,且该基因与小麦抗旱性密切相关。秦鹏等[22]研究指出,在干旱处理下敏感型小麦叶片TaWdreb2、TaWlip19基因表达显著低于抗旱型小麦,说明小麦受到干旱胁迫后不同品种的基因表达存在差异,这与小麦品种自身抗旱能力和抗旱调控途径具有一定的相关性。
综上所述,目前对干旱胁迫的研究主要集中在植株光合特征和生理生化指标动态变化规律上,或分子水平基因表达的单因素研究上,而关于干旱胁迫条件下春小麦开花期光合特性、生理生化指标以及根系抗旱基因表达的系统性研究报道较少,尤其是对于多指标间协同效应的研究更少。本研究以抗旱组3个耐旱春小麦品种(定西40号、龙麦36号、龙麦33号)和水敏感组3个水敏感春小麦品种(农麦2号、巴麦12号、巴丰5号)为研究对象,在大兴安岭西麓旱作区内蒙古自治区农牧业科学院特泥河土壤管理与生态修复科学观测试验站,设置旱棚防雨和调控补水2个水分控制处理,系统分析各春小麦品种的旗叶生理特征及根系抗旱基因表达差异,明确水敏感组和抗旱组春小麦的各指标差异性,为揭示耐旱春小麦对干旱胁迫的适应机制及抗旱栽培提供了理论依据和实践基础。
1 材料与方法
1.1 供试材料
根据国家现代产业技术体系春小麦抗旱品种的推荐,结合查询前人研究结果和课题组内部的春小麦抗旱性试验验证,筛选出龙麦36号(LM36)、龙麦33号(LM33)、定西40号(DX40)3个耐旱春小麦品种和农麦2号(NM2)、巴麦12号(BM12)、巴丰5号(BF5)3个水敏感品种作为供试材料。
1.2 试验地概况
试验于2019年在内蒙古自治区农牧业科学院特泥河土壤管理与生态修复科学观测试验站(120°48′E,49°55′N,海拔650 m)开展。试验区地处中温带半干旱大陆性草原气候区,无霜期90~105 d,年平均日照时数2 589 h,地表土质为黑钙土,属于典型内蒙古农牧交错带大兴安岭西麓黑土地代表。土壤养分:碱解氮110~120 mg·kg-1、速效钾144 mg·kg-1、速效磷20 mg·kg-1,pH 6.8。2000—2019年平均降水量为334.86 mm,2019年特泥河年降水量330.2 mm,2019年春小麦全生育期降雨量为223.9 mm(图1)。在春小麦全生育期,7月降雨量最大为92.0 mm,占全年降水量的27.86%。

注:图中虚线框为旱棚防雨-调控补水的水分控制处理时间,为7月5日至7月25日。
1.3 试验设计
试验采用裂区设置,其中,主处理为水分处理,包括干旱(HC)处理和对照(CK)处理;副处理为春小麦品种,主要包括抗旱组(DRG)3个品种(DX40、LM36、LM33),水敏感组(WSG)3个品种(NM2、BM12、BF5)共6个品种处理,共12个处理,每个处理重复3次,共36个小区,每个小区面积为9 m2,区组间距为0.5 m,设置1 m宽保护行。
基于前期试验结果,春小麦水敏感品种比耐旱品种的开花期提前15 d左右。因此,为保证6个春小麦品种的水分处理在同一生育期内,2019年抗旱春小麦品种DX40、LM36、LM33的播种日期为5月5日,水敏感组春小麦品种NM2、BM12、BF5的播种日期为5月20日。播种量均为300 kg·hm-2,行距为0.15 m。播种时所有肥料一次性施入,其中尿素施入量为60 kg·hm-2、磷酸二铵施入量为180 kg·hm-2、硫酸钾施入量为30 kg·hm-2,后期不追肥,除水分处理外,其他管理方式同大田。干旱处理在旱棚内进行,将土壤质量含水量控制在8%~12%;对照组根据春小麦生长发育所需水量,通过雨水自养与漫灌相结合的方式将土壤质量含水量控制在25%~30%之间。干旱处理时间为22 d,于春小麦拔节期(7月4日)开始到春小麦开花后期(7月25日)结束。在水分控制处理的22 d内,试验共进行5次土壤质量含水量监测,监测具体时间为2019年7月4日、7月10日、7月15日、7月22日、7月26日。通过监测干旱处理最终土壤质量含水量为10.83%,对照处理最终土壤质量含水量为29.12%,均在试验预设范围内(图2)。
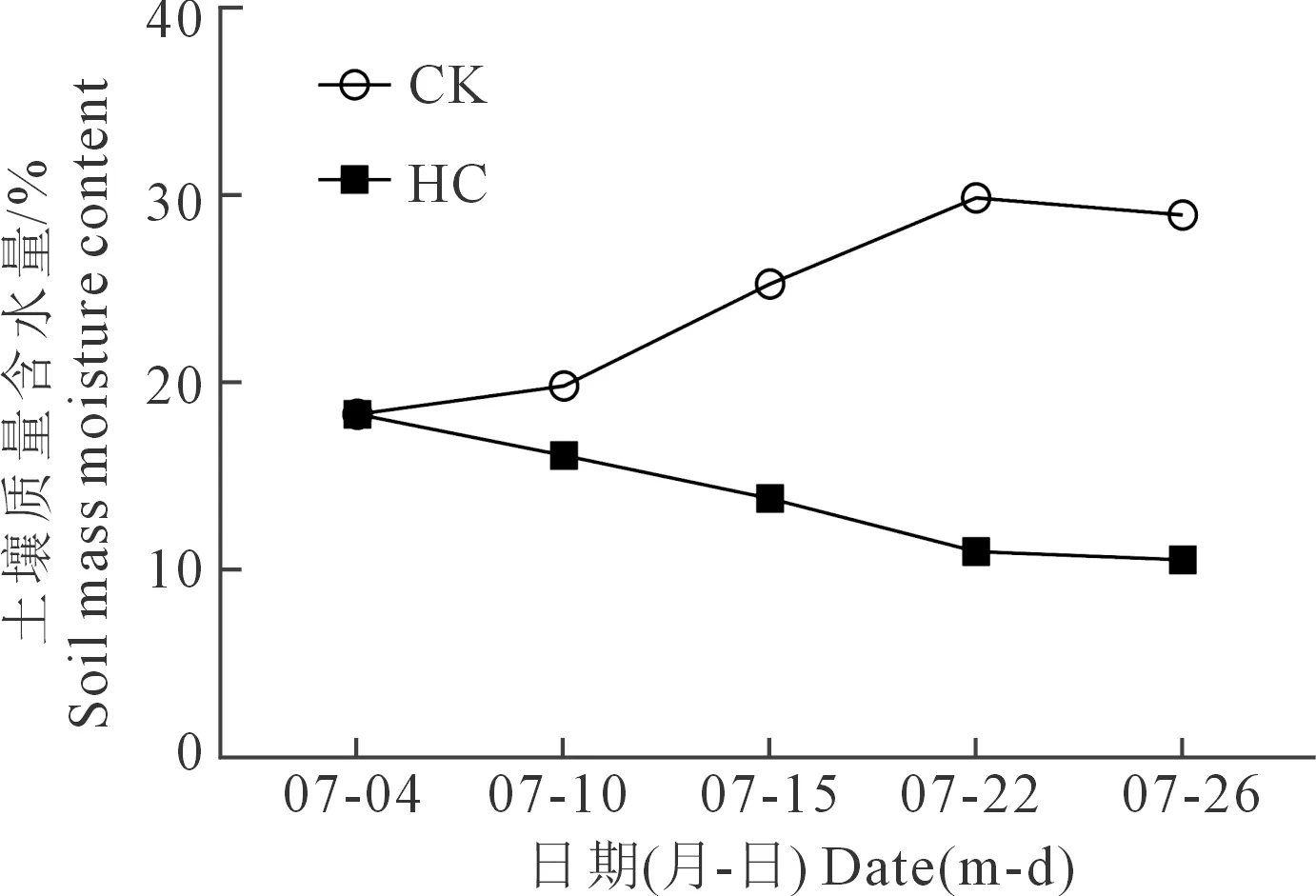
注:不同处理土壤质量含水量变化情况,对照处理(CK)共灌水两次(7月11日、7月16日),灌水量为:每个处理3次重复共2 m3。
1.4 测定指标及方法
1.4.1 春小麦开花期旗叶光合特性指标测定 在春小麦开花后期(全田90%以上麦穗中上部小花的内外颖张开,花药散粉且部分籽粒出现灌浆)选择无风无云的晴天进行测定,每小区选取长势均匀的3株春小麦测定旗叶光合特性及SPAD,重复3次。
光合特性:采用便捷式光合仪(LI-6800,LI-COR公司,美国),在9∶00—11∶30将红蓝光源设定为1 200 μmol·m-2·s-1,测定春小麦旗叶净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)和蒸腾速率(Tr)。
SPAD:采用叶绿素测定仪(TYS-A,浙江拓普仪器有限公司)将春小麦旗叶(该叶片与测定光合特性叶片相同)放入叶室内并将叶室完全覆盖,将上下叶室夹紧后读数,每个叶片重复读数3次,取平均值作为该植株叶片的SPAD。
1.4.2 春小麦开花期旗叶生理指标测定 在每个小区内选取长势均匀且具有代表性的春小麦9株,用无菌剪刀将春小麦旗叶剪下,无菌水冲洗干净叶片表面的附着物,用吸水纸吸干叶片表面残留的无菌水,锡箔纸包裹样品,做好标记,液氮速冻后放于-80℃超低温冰箱保存备用。过氧化物酶(POD)活性、超氧化物歧化酶(SOD)活性、丙二醛(MDA)含量、脯氨酸(Pro)含量均参照高俊凤[23]的方法测定。
1.4.3 春小麦地上部干重指标测定 春小麦开花后期,在每个小区内选取长势均匀且具有代表性的春小麦5株,取其地上部称重,重复3次。将称完鲜重的春小麦放于烘箱内105℃杀青0.5 h后,80℃烘干至恒重,称重。
1.4.4 春小麦开花期根系样品采集及保存 在每个小区内选取长势相同且具有代表性的春小麦9株,选取新鲜幼嫩生长旺盛的根,用无菌水快速清洗根表面的附着物,用吸水纸吸干根系表面残留的无菌水,用无菌剪刀剪成1 cm的小段。将3次重复的春小麦样品等量混合均匀后,放置于3个5 mL灭菌的离心管中,做好标记液氮速冻后放于-80℃超低温冰箱保存备用,样品用于总RNA的提取。
1.4.5 引物的设计与合成 参考GenBank(https://www.ncbi.nlm.nih.gov/genbank/)中的XTH-7A、Wlip19、Wdreb2、BADHb、Actin(实时荧光定量内参基因)基因cDNA序列(GenBank登录号分别为:MK395550、AB193552.1、AB193608.1、AY050316.1、AK458303.1),利用Primer5.0软件设计引物,用于普通PCR和实时荧光定量PCR(RT-qPCR)[24]。引物由北京盛元科萌基因生物科技有限公司合成,引物序列信息见表1。

表1 引物序列信息
1.4.6 总RNA提取质量检测及反转录 参照Easy Pure Plant RNA Kit试剂盒(目录号ER301,北京全式金生物技术有限公司)。用1%的琼脂糖凝胶以及超微量核酸定量仪(NanoDrop 2000,美国)检测提取RNA的浓度及质量。记录浓度(ng·μL-1)及A260/A280比值。参照TransScript All-in-One First-Strand cDNA Syn One-Step gDNA Removal thesis SuperMix for qPCR(One-Step gDNA Removal)试剂盒(目录号AT341,北京全式金生物技术有限公司)。20 μL反应体系为Total RNA/ mRNA 800 ng,5×TransScript®All-in-One SuperMix for qPCR 4 μL,gDNA remover 1 μL,RNase-free Water Variable。设置PCR反应程序为:(42℃ 15 min)×1;(85℃ 5 s)×1;(4℃ ∞)。将反转录得到的cDNA(质量浓度为1 μg·μL-1)放置于-80℃冰箱保存。
1.4.7 目的基因的PCR扩增 25 μL的PCR体系为cDNA(1 μg·μL-1) 2 μL,2×EasyTaq® PCR SuperMix 12.5 μL,上、下游引物(10 μmol·L-1)各1 μL,Nuclease-free Water 8.5 μL。PCR反应条件为:(94℃ 5 min)×1;(94℃ 30 s;58℃ 30 s;72℃ 30 s)×35;(72℃ 10 min)×1;(4℃ ∞)。
1.4.8 实时荧光定量PCR(RT-qPCR) 参照TransStart Tip Green qPCR SuperMix试剂盒(目录号AQ601,北京全式金生物技术有限公司)。荧光定量PCR反应体系为cDNA(1 μg·μL-1)1 μL,2×PerfectStartTMGreen qPCR SuperMix 10 μL,上、下游引物(10 μmol·L-1)各0.8 μL,Nuclease-free Water 7.4 μL。利用荧光定量PCR仪(Light Cycler 4800Ⅱ)进行qRT-PCR扩增,反应条件:95℃预变性30 s;95℃变性5 s;60℃退火30 s;40个循环。每个样品设置3个生物学重复,3个技术重复。
1.5 统计分析
春小麦根系抗旱基因表达水平采用2-ΔΔCt的计算方法[19],ΔCt=Cttarget gene-Ctactin。
采用Excel 2010和IBM SPSS Statistics 22软件进行春小麦旗叶光合特性、生理指标和根系抗旱基因表达试验数据计算及统计分析,采用单因素方差分析(ANOVA),新复极差法(Duncan)的多重比较方法进行差异显著性检验(α=0.05)。当P<0.05时具有显著性差异;P<0.01时具有极显著差异;P>0.05时则无显著性差异。对春小麦旗叶各指标进行Spearman相关性分析。利用GraphPad Prism 8作图。
2 结果与分析
2.1 干旱胁迫对耐旱春小麦旗叶光合特性的影响
与对照相比,干旱处理显著降低了春小麦旗叶净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、SPAD(P<0.05)(图3);而胞间CO2浓度(Ci)则呈相反结果,干旱处理下春小麦旗叶Ci显著增加(P<0.05)。其中抗旱组春小麦旗叶Pn降低15.74%;Gs下降40.74%;Tr下降34.32%;相对叶绿素含量(SPAD)降低7.58%;Ci提高7.77%,而水敏感组Pn降低35.15%;Gs下降48.98%;Tr下降39.29%;SPAD降低13.56%;Ci提高9.09%(图3A~E)。表明干旱处理会降低春小麦旗叶Pn、Gs、Tr、SPAD,增加Ci,而抗旱组Pn、Gs、Tr、SPAD下降幅度和Ci的上升幅度明显小于水敏感组。
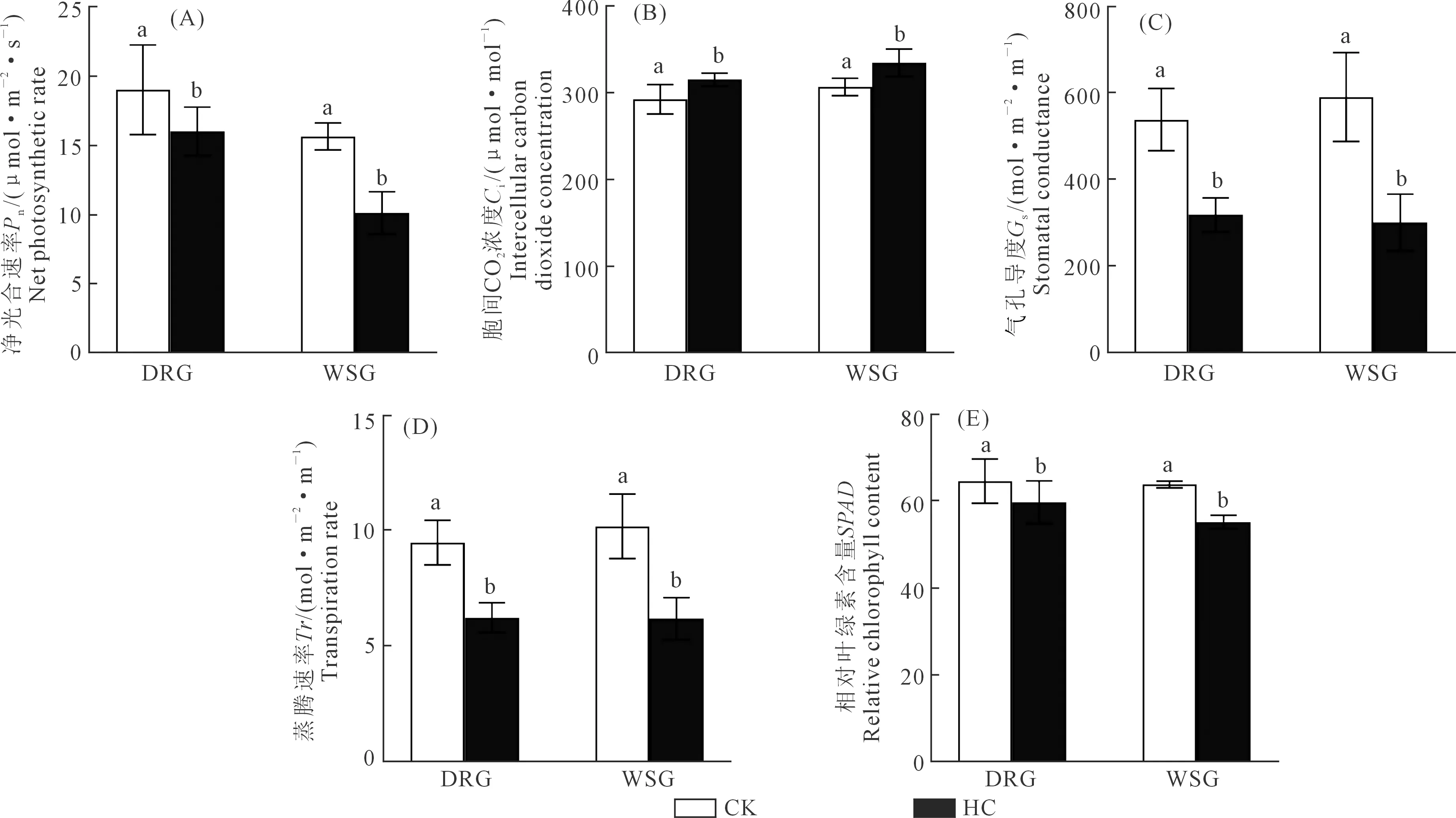
注:(A)净光合速率;(B)胞间CO2浓度;(C)气孔导度;(D)蒸腾速率;(E)SPAD;DRG:抗旱组(DX40、LM36、LM33三个品种的均值);WSG:水敏感组(NM2、BM12、BF5三个品种的均值)。图上不同小写字母表示同一组别春小麦在不同处理间P<0.05水平下显著,下同。
干旱处理下春小麦抗旱组与水敏感组相比,具有较高的Pn、Gs、Tr和SPAD。从光合特性的角度来看,抗旱组春小麦叶片持绿性较好、衰老较慢,叶片水、气循环与物质交换条件良好,利于其干物质的积累。因此,抗旱组春小麦表现出较强的抗旱性,更能适应干旱环境。
2.2 干旱胁迫对耐旱春小麦旗叶生理指标的影响
与对照相比,干旱处理下春小麦旗叶过氧化物酶(POD)、超氧化物歧化酶(SOD)活性、丙二醛(MDA)、脯氨酸(Pro)含量显著增加(P<0.05)(图4);抗旱组分别提高了14.18%、2.00%、29.78%,而水敏感组分别提高了17.06%、7.38%、73.80%;其中抗旱组春小麦旗叶Pro含量为90.36 μg·g-1,水敏感组为70.36 μg·g-1;水敏感组春小麦旗叶MDA含量比抗旱组提高11.24%(图4A~D)。表明抗旱组生理指标总体变化幅度小于水敏感组,且抗旱组Pro含量明显高于水敏感组,MDA含量明显低于水敏感组。

注:(A)过氧化物酶活性;(B)超氧化物歧化酶活性;(C)丙二醛含量;(D)脯氨酸含量;DRG:抗旱组(DX40、LM36、LM33三个品种的均值);WSG:水敏感组(NM2、BM12、BF5三个品种的均值)。
综合比较,抗旱组春小麦旗叶Pro含量比水敏感组高,而POD、SOD活性、MDA含量比水敏感组低。从植株生理指标的角度来看,抗旱组较水敏感组春小麦旗叶细胞膜脂损伤程度低、叶片组织失水少、抗氧化能力强,所以抗旱组春小麦能较好地维持春小麦的正常生理功能,抵抗干旱危害的能力较强。
2.3 干旱胁迫对耐旱春小麦根系抗旱基因表达特征的影响
干旱胁迫下,作物光合生理及植株生理指标均会发生显著变化。而不同春小麦品种其基因型不同。基于文献资料[19-21]发现,在小麦受到干旱胁迫时TaXTH-7A、TaBADHb、TaWdreb2、TaWlip19等基因会发生显著变化。
本研究中抗旱组春小麦根系基因TaXTH-7A、TaWlip19、TaWdreb2、TaBADHb相对表达量表现为干旱处理显著高于对照处理,水敏感组根系基因TaXTH-7A、TaWlip19、TaWdreb2、TaBADHb相对表达量则为干旱处理显著低于对照处理(P<0.05)(图5A~D)。干旱处理下抗旱组春小麦根系基因TaXTH-7A、TaWlip19、TaWdreb2、TaBADHb相对表达量分别比对照显著提高1.067倍、1.737倍、1.036倍、0.799倍。而水敏感组春小麦根系基因TaXTH-7A、TaWlip19、TaWdreb2、TaBADHb相对表达量分别比对照显著降低了0.735倍、0.554倍、0.667倍、0.496倍(图5A~D)。

注:(A)TaXTH-7A相对表达量;(B)TaWlip19相对表达量;(C)TaWdreb2相对表达量;(D)TaBADHb相对表达量;DRG:抗旱组(DX40、LM36、LM33三个品种的均值);WSG:水敏感组(NM2、BM12、BF5三个品种的均值)。
综上,干旱处理下TaXTH-7A、TaWlip19、TaWdreb2、TaBADHb基因在抗旱组春小麦根系中相对表达量均显著上调,而在水敏感组中4个基因表达均显著下调,从抗旱基因表达水平上反映出抗旱组春小麦可通过上调抗旱基因表达应对干旱危害。
2.4 干旱胁迫对耐旱春小麦地上部干重的影响
与对照相比,干旱处理下春小麦干重均显著降低(P<0.05),抗旱组春小麦干重降低了27.77%,而水敏感组降低了32.39%,表明干旱处理会显著降低春小麦干重,抗旱组春小麦可以保持相对较好的生长状况,积累更多的干物质(图6)。

图6 干旱处理对抗旱组、水敏感组春小麦干重的影响
2.5 春小麦生物量与春小麦旗叶生理特征的相关分析
为了明确春小麦生物量与春小麦旗叶生理特征之间的相关关系,本试验以春小麦地上部干重作为其生物量的代表与上述各指标进行了Spearman相关性分析,结果见表2。

表2 春小麦地上部干重与春小麦旗叶生理特征相关系数
春小麦地上部干重与光合指标呈显著正相关的有4个:相对叶绿素含量(X1)、蒸腾速率(X2)、净光合速率(X3)、气孔导度(X4);相关系数分别为0.549、0.604、0.585、0.605。春小麦地上部干重与胞间CO2浓度(X5)呈显著负相关,相关系数为-0.585。
春小麦地上部干重与生理指标呈显著负相关的有2个:超氧化物歧化酶(X7)、丙二醛(X8),相关系数分别为-0.356、-0.440。
3 讨 论
3.1 耐旱春小麦通过调节叶片水、气循环条件适应干旱逆境
光合作用是植物重要的生理过程之一,也是植物生长发育和产量形成的基础,还是外部生态因子和内部生理因子共同作用的复杂过程[25]。本试验在光合特性方面的研究表明,干旱处理下抗旱组和水敏感组春小麦旗叶SPAD、Pn、Tr、Gs显著降低,抗旱组分别降低了7.58%、15.74%、34.32%、40.74%,而水敏感组分别降低了13.56%、35.15%、39.29%、48.98%;Ci显著升高,抗旱组提高7.77%,水敏感组提高9.09%,抗旱组各光合指标变化幅度小于水敏感组。本研究结果在前人的研究中也得到了印证[26-28],并得出相同结论,即干旱胁迫会导致春小麦旗叶持绿性差,叶片衰老加快,不利于叶片细胞物质交换,光合作用强度减少,影响春小麦生长发育。
一定条件下,气孔导度(Gs)和胞间CO2浓度(Ci)呈正相关关系[12],在干旱和对照两种水分处理下抗旱组春小麦旗叶SPAD、Pn、Tr、Gs下降幅度均小于水敏感组,干旱胁迫对抗旱性强的品种光合作用影响要比水敏感品种低且抗旱性强的品种可以保持相对较高的Gs,为Pn和Tr提供有利条件,而较高的Pn和Tr可以促进作物积累较多的干物质,促使抗旱强的作物保持较高的产量[29]。Ci的上升幅度小于水敏感组,抗旱组Ci上升幅度较小也说明了Pn下降是由于非气孔因素导致的,这与张士昌、张东等[11,28]的研究结果相同。本研究还发现相对叶绿素含量、蒸腾速率、净光合速率、气孔导度与干重呈显著正相关。说明以上指标与干物质积累程度密切相关,可通过测定春小麦旗叶光合参数:SPAD、Pn、Tr、Gs等的变化趋势,评估干旱对春小麦生育后期干物质积累的危害程度,进而量化干旱对作物造成的影响。
3.2 耐旱春小麦通过增强抗氧化酶活性和水溶性物质含量缓解干旱损伤
作物在正常生长发育阶段会产生活性氧自由基(ROS),尽管ROS有可能引起有害的氧化,但它也是一种参与控制植物生长发育的信号分子[30],在干旱胁迫下,光抑制和光呼吸会导致大量的ROS积累,对作物的细胞膜系统造成伤害,但作物本身会启动相应的机制来激活细胞防御系统进而抵抗逆境危害[31]。本试验通过对春小麦旗叶生理指标的研究发现:干旱胁迫下抗旱组、水敏感组春小麦旗叶POD、SOD、MDA、Pro均显著升高。在干旱处理下水敏感组春小麦旗叶POD、SOD、Pro上升幅度均显著高于抗旱组。这说明在干旱条件下,植物细胞内清除活性氧的酶促体系(SOD、POD等)的含量有升高的趋势,能够帮助清除植物机体内的活性氧有害物质以提高植物的抗旱性[32-33];孙军伟等[34]研究表明,在小麦灌浆期遭受干旱胁迫时Pro含量急剧上升,在一定程度上减轻叶片组织失水,较好地保持了植株的正常生理功能[11,35],而抗旱性强的品种Pro含量显著高于抗旱性弱的品种,渗透物质的增加可以使抗旱性强的品种保持相对较好的水分平衡。干旱处理下,抗旱性强的品种其MDA含量上升较小,生物膜损伤程度较小[36-38]。这与本研究结果一致。
3.3 耐旱春小麦通过上调抗旱基因表达抵御干旱危害
小麦基因的复杂多样性可能促使小麦在干旱的环境下生长[16],这与小麦基因的表达差异有关。在基因表达方面,我们基于前人的研究结果,在抗旱功能基因和抗旱调节基因方面分别选取了2对典型的抗旱相关基因(XTH-7A、Wlip19,Wdreb2、BADHb)作为对基因表达分析的代表。杨成民等[39]研究表明,XTH-7A、BADHb基因在植物抗逆过程中编辑产生功能蛋白,直接对植物细胞起到保护作用。周琪等[20]研究发现,在干旱胁迫下TaXTH-7A有利于作物提高渗透调节能力和更好地存活。李素等[21]研究表明,TaBADHb基因表达呈上调趋势且该基因与小麦抗旱性密切相关。刘欣等[40]研究表明,Wdreb2、Wlip19基因在植物抗逆过程中传导信号及参与抗旱相关基因表达的调控。秦鹏等[22]研究指出,敏感型小麦叶片在干旱胁迫下的TaWdreb2、TaWlip19基因表达显著低于抗旱性小麦,说明基因在不同品种小麦受到干旱胁迫后存在表达差异,这与小麦品种的抗旱能力具有一定相关性。在干旱处理下XTH-7A、Wlip19、Wdreb2、BADHb基因在抗旱组中均为高表达,在水敏感组中均为低表达,抗旱组4种基因相对表达量比对照分别显著提高了1.067倍、1.737倍、1.036倍、0.799倍,与前人研究结果一致。
另一个有趣的发现是,除XTH-7A基因外,前人主要在小麦叶片上对Wlip19、Wdreb2、BADHb进行基因表达差异的研究,而本试验发现在根系中也可得到与前人相似的结果。由于植物不同的组织细胞生活的环境不同,基因表达具有选择性,因此经过转录产生的mRNA不完全相同。因此,本试验结果也可为以后的研究提供参考。
4 结 论
干旱胁迫下春小麦通过关闭或部分关闭气孔降低气孔导度,减少水分蒸发。光抑制和光呼吸会产生大量ROS积累,对作物细胞膜系统造成伤害。抗旱组春小麦积极上调根系抗氧化酶类基因、渗透调节等相关基因表达来提高其旗叶POD、SOD等抗氧化酶活性和增加Pro等水溶性物质在胞质中的含量,降低细胞内活性氧浓度,减少其对细胞的毒害作用,从而缓解了干旱对春小麦生长的抑制作用。耐旱春小麦通过减小根系基因表达与旗叶的光合特性及生理指标的变化幅度,以此增强其干旱适应性。抗旱组春小麦提高抗旱性能并促使其较水敏感组春小麦更能抵御干旱胁迫的危害。因此,抗旱组春小麦的抗旱性较好。
