甘肃天水市退耕还林区土壤纤毛虫群落多样性
2022-05-16宁应之沈小亚张惠茹杨怀印陈凌云
宁应之,沈小亚,张惠茹,杨怀印,陈凌云
(西北师范大学生命科学学院,甘肃兰州 730070)
退耕还林工程是我国投资巨大、涉及面极广、利在当代、泽及千秋的重大生态工程,对于我国、尤其是我国生态环境极其脆弱的西北地区的植被恢复、生态环境改善以及经济和社会的可持续发展具有极其重要的意义.甘肃是我国首批实施退耕还林工程的3个试点省份之一,自1999年至今已实施退耕还林工程22年.退耕还林工程的实施,不仅优化了农村和牧区产业结构,增加了农民和牧民收入,促进了农村和牧区的建设和发展,更重要的是增强了全民环境意识,提高了植被覆盖率,改善了区域生态环境.退耕还林导致地上生态环境发生的变化必然影响到地下,使土壤环境质量发生变化,而这又必然引起土壤动物群落在组成、结构和功能上的响应,环境敏感动物群落的响应会更为积极而显著——土壤纤毛虫群落正是这种环境敏感动物群落.土壤纤毛虫是土壤原生动物重要类群之一,泛指栖息于土壤孔隙水中、土壤颗粒和凋落物表面水膜中的纤毛虫[1],其物种繁多、现存量和生产量巨大、个体微小、细胞膜纤薄、生长繁殖快、群落演替迅速、分布广泛,因而成为对生态变化反应最为敏感的动物类群,可广泛、快速、有效和经济地用于反映、评价、监测甚至预报土壤生态环境的变化[2].
如前所述,由于纤毛虫在响应生态变化方面具有极大的优势,所以从上世纪20年代起开始研究土壤纤毛虫群落(及原生动物其他类群)对土壤生态变化的响应并据此评价和监测土壤生态变化[3].但这些研究工作绝大多数聚焦于土壤纤毛虫群落(及原生动物其他类群)对人类恶性干扰导致土壤生态恶化的响应并通过响应参数来评价和监测生态恶化状况[4-19].近年来,随着生态恢复进程的加速,国内陆续有对生态恢复区土壤动物的研究,其中包括土壤动物对退耕还林生态恢复响应的研究[20-23],但这些工作都只涉及中、小型动物类群,有关土壤原生动物(包括土壤纤毛虫)的工作较少,仅有宁应之等对甘肃部分地区土壤纤毛虫群落对于退耕还林生态恢复的响应研究以及基于研究结果对退耕还林生态恢复效果进行评价的研究工作,研究区域较为零散,未形成一个范围较大的退耕还林整体区域内的系统性研究工作[24-28].
为探讨退耕还林不同典型林地对土壤纤毛虫群落多样性的影响,文中对天水市麦积区境内退耕还林区土壤纤毛虫群落多样性的现状进行研究,以期为该地区退耕还林工程的生物学评价体系提供微型生物基础资料.
1 研究区概况
研究区位于天水市麦积区伯阳镇,地理坐标为105°25′~106°43′ E,34°06′~34°48′ N,东接陕西省宝鸡市,西邻甘肃甘谷县,属暖温带半干旱大陆性季风气候,年降雨量从南向北依次减少,每年560 mm左右,年平均气温为10.9 ℃,全年光照2 800 h,年有效积温3 000 ℃以上,无霜期近200 d[29].植被优势种为野菊(Dendranthemaindicum)、狗尾草(Setariaviridis)、艾蒿(Artemisiaargyi)和白藜(Chenopodiumalbum)等.当地黄绵土、黑垆土为主要耕作土壤.
2 材料与方法
2.1 野外工作
2.1.1 土壤样品采集 在退耕区选择退耕种植刺槐的纯种生态林(CH),种植苹果的纯种经济林(PG)和种植花椒+核桃的混交经济林(HJHT)3种典型生态恢复林区作为实验样区,以耕地(GD)作为对照样区.本研究选择的实验样区均为退耕还林17年的林地.
采样按照季节分4次进行,分别是2016年6月(夏)、9月(秋)、11月(冬)和2017年3月(春).每个采样点设置5组面积为25 m2(5 m×5 m)的平行样方,采样时除去土层表面凋落物,采用梅花五点式取样法用采样器(15 mL圆筒形)采集0~5 cm土壤表层土样,标记并保存,共计20份[25].
2.1.2 相关环境因子测定 采样时,用土壤环刀取样器(100 cm3)切割原位土样,连同取样器装袋标记并保存,用于测定土壤孔隙度、土壤容重等指标,同时用曲管地温计测量土壤温度(测量温度范围:-10~60 ℃),记录样区海拔、地理坐标,观察土壤质地.
2.2 室内工作方法
采集到的土壤样品需分开处理,一部分新鲜土样测定含水量和pH值,剩余土样置于通风处自然风干后装袋保存,用于土壤纤毛虫的定性和定量研究,风干过程中在土样表面覆盖一层草纸,防止外来土壤纤毛虫包囊污染样品[25].
2.2.1 纤毛虫定性和定量研究 称取50 g土壤样品,置于直径为15 cm的培养皿中,加入土壤浸出液充分浸润但不淹没土壤,此方法为非淹没培养皿法[30],培养的土样用于做定性研究.将培养皿置于25 ℃的恒温光照培养箱中培养,24 h后开始镜检,连续培养观察20 d,并持续做好活体观察记录,期间需定期添加土壤浸出液,使土壤样品处于“非淹没”状态,同时结合Wilbert蛋白银染色法进行鉴定[31],物种鉴定所用资料见文献[32-35],最后依据Lynn(2008)分类系统[36]进行分类整理.
定量研究采用培养直接计数法,风干土壤样品和蒸馏水按照1∶1的比例进行配置.称取30 g土壤样品置于培养皿中,加入30 mL水,记录培养皿液面,随后将其置于25 ℃的恒温光照培养箱中培养,维持液面不变直至最大计数日(第9,10,11 d),在最大计数日进行实验.将培养皿倾斜45°,静置5~7 min,转移全部土壤上清液并计量,从中吸取一滴于显微镜下计录纤毛虫总数及个体数量,1份土样重复观察5次。将所得结果换算为1 mL(22滴约等于1 mL)水中的纤毛虫总数,最后可换算为30 g土壤样品中纤毛虫的密度.
2.2.2 土壤理化因子测定 土壤含水量采用烘干法测定参照文献[37];土壤孔隙度和土壤容重用环刀法测定参照文献[38];土壤pH值采用电位法测定(水∶土=2.5∶1);土壤电导率采用浸提法测定(水∶土=5∶1).兰州大学草地农业科技学院草地生态化学计量学实验室测定并提供土壤总磷、总钾、总氮和有机质数据.
2.2.3 数据统计分析 统计各样点纤毛虫物种分布情况并进行类群划分,在目级分类阶元上,将物种数占土壤纤毛虫物种总数比例最大的目定义为优势类群,次大的目定义为次优势类群,仅有1种的目定义为罕见类群,其余各目为常见类群[25].
相关指数和系数计算如下:
C/P系数:C/P=肾形纲纤毛虫物种数/瓶纤纲、异毛纲、旋毛纲纤毛虫物种数.
Shannon-Wiener指数(H):
Simpson指数(C):
Pielou指数(E):
Margalef指数(D):
其中,N为群落中所有属的个体总数;Pi为每个样点中第i个属土壤纤毛虫的个体密度占该样点土壤纤毛虫总数的比例;S为所有种类数.
采用SPSS 23.0统计分析软件对数据进行统计分析(One-way ANOVA分析,平均数采用LSD进行多重比较分析).应用Arcview gis 10.2绘制采样区地图;Primer 6.0进行BIOENV相关性分析和聚类分析;Origin 2018绘制柱状图和折线图.
3 结果与分析
3.1 纤毛虫群落多样性特征
3.1.1 群落组成 共鉴定到土壤纤毛虫139种,隶属于9纲20目39科55属,散毛目(Sporadotrichida)为优势类群,占总物种数的21.58%;刺钩目(Haptorida)和肾形目(Colpodida)为次优势类群,分别占总物种数的18.71%和12.23%;喙纤目(Loxodida)、原口目(Protostomatida)、弹跳目(Halteriida)、咽膜目(Peniculida)为罕见类群,合计占总物种数的2.88%;异毛目(Heterotrichida)、游仆目(Euplotida)、排毛目(Stichotrichida)等13个目为常见类群,合计占总物种数的44.60%.随着退耕年限的增加,3个林地间纤毛虫物种数和优势类群差异显著(表1),刺槐林物种最丰富,分布有93种,占物种总数的66.90%,优势类群为散毛目(Sporadotrichida),占该样点物种总数的30.00%;花椒核桃林次之,分布有85种,占物种总数的61.15%,优势类群为散毛目(Sporadotrichida),占该样点物种总数的22.00%;苹果林和耕地分别分布有80和52种,占物种总数的57.55%和37.41%,优势类群分别为刺钩目(Haptorida)、肾形目(Colpodida),占该样点物种总数的25.00%和21.00%.从优势类群的演替趋势来看,总体呈现为耕地较为低等的类群肾形目(Colpodida)演替为退耕后林地较为高等的类群散毛目(Sporadotrichida).可见退耕后林地土壤纤毛虫的群落组成与耕地差异显著,退耕样点的群落组成更为复杂、物种数更多.
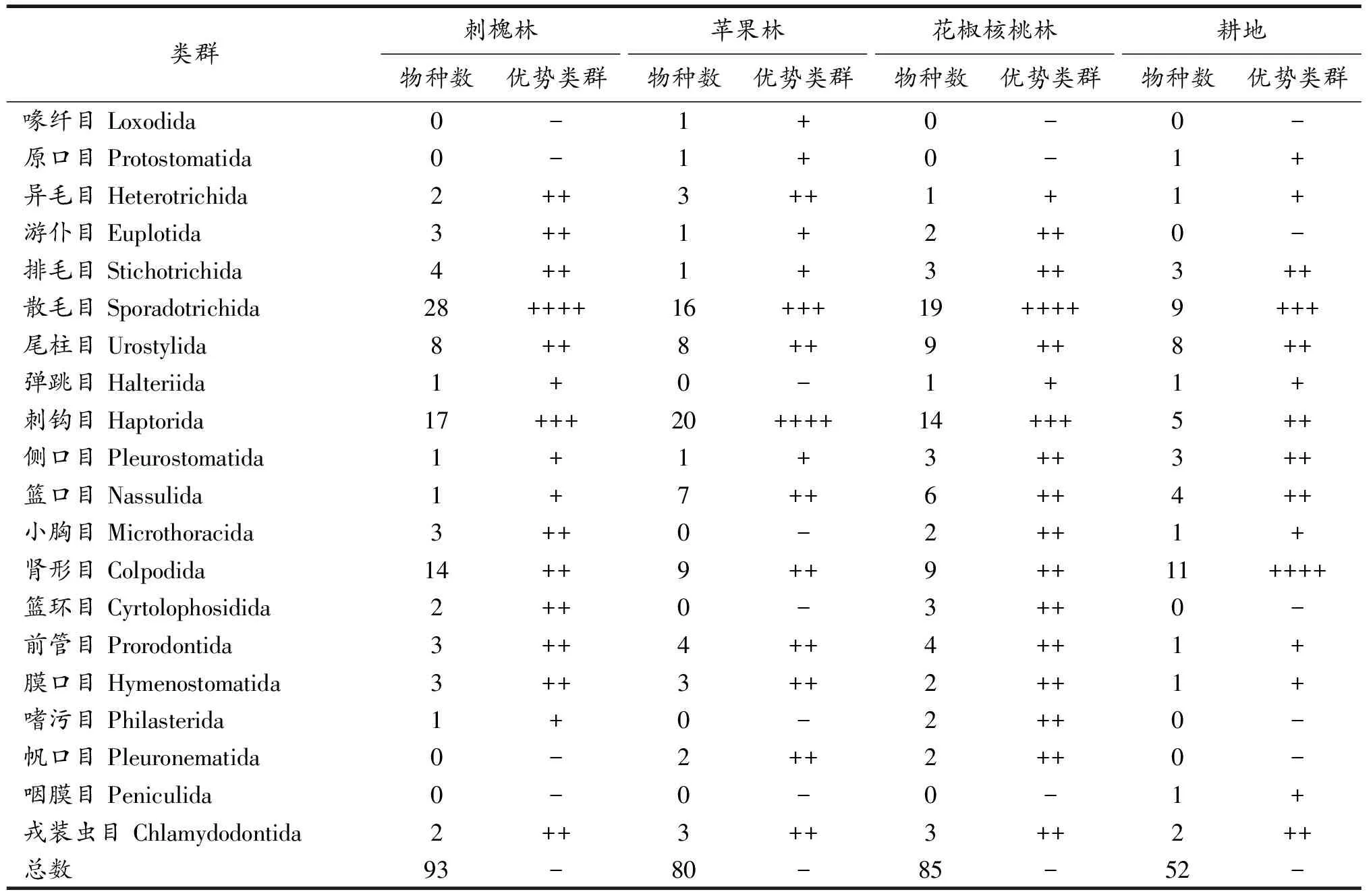
表1 各样点土壤纤毛虫群落组成
图1为各样点土壤纤毛虫群落特征参数,结果显示,耕地的土壤纤毛虫物种数在四个季节中均最少,分别为19(春)、18(夏)、27(秋)、17(冬);刺槐林物种数均最多,分别为46(春)、37(夏)、45(秋)、35(冬)(图1(a));四季的平均密度表现为退耕样点大于耕地且差异显著(P<0.05),总体呈现刺槐林(122.72 ind·g-1)>花椒核桃林(120.05 ind·g-1)>苹果林(84.75 ind·g-1)>耕地(38.65 ind·g-1)的趋势(图1(b));耕地的C/P系数在四季中均大于0.8,且显著高于退耕样点(P<0.05).即低等类群肾形纲数量较多,土壤纤毛虫群落结构简单,而退耕样点的C/P系数在0.17~0.58之间波动,说明肾形纲土壤纤毛虫的数量减少,异毛纲和旋毛纲土壤纤毛虫的数量增多(图1(c)).
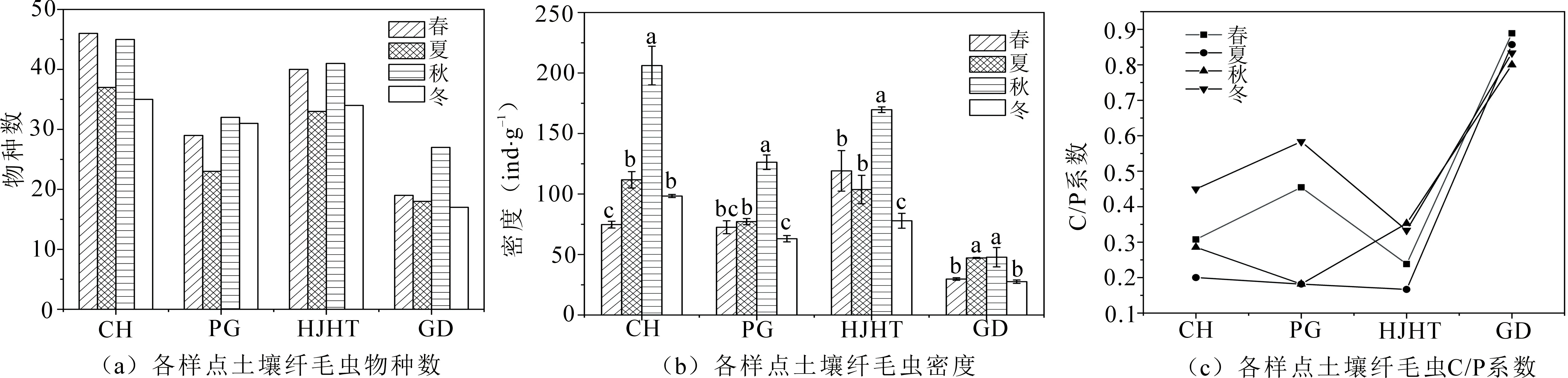
图1 各样点土壤纤毛虫群落特征参数
3.1.2 群落多样性指数 图2为各样地土壤纤毛虫群落多样性指数,总体表现出耕地土壤纤毛虫Shannon-Wiener指数(H=1.84)、Simpson指数(C=0.75)、Margalef指数(D=2.90)和Pielou指数(E=0.75)都显著低于退耕样点(P<0.05),退耕样点中,Shannon-Wiener指数和Margalef指数均表现为刺槐林最高(H=2.40,D=4.13),花椒核桃林次之(H=2.27,D=3.80),苹果林最低(H=2.18,D=3.38)的趋势(图2(b)-(c)),而Simpson指数和Pielou指数则表现为刺槐林最高(C=0.84,E=0.80),苹果林次之(C=0.82,E=0.79),花椒核桃林最低(C=0.81,E=0.77)的趋势(图2(a)-(d)).由此可见,退耕还林后林地土壤纤毛虫群落多样性指数增大,表明林地土壤纤毛虫群落结构趋于稳定,物种趋于复杂.
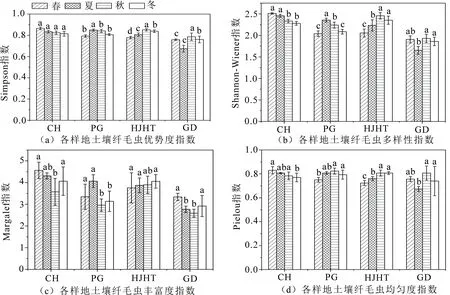
图2 各样点土壤纤毛虫多样性指数
3.1.3 群落相似性 对4个样点土壤纤毛虫群落特征参数进行聚类分析(图3),结果显示,花椒核桃林与刺槐林最先在相似性为98的水平上聚为一类,然后与苹果林在相似性为95的水平上聚为一类,最后与耕地样点在相似性为84的水平上聚为一类.表明退耕还林后,刺槐林与花椒核桃林土壤纤毛虫群落相似性水平最高,与苹果林也较为相似,但这3块样地与耕地的相似性较低.因此,退耕还林后林地中土壤纤毛虫群落结构更相似,与耕地差异较大.
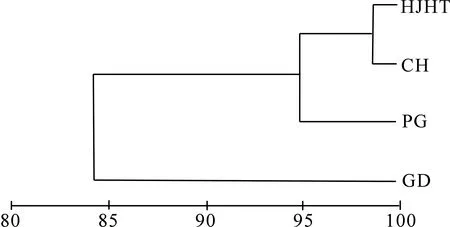
图3 各样点土壤纤毛虫群落的聚类分析
3.2 纤毛虫群落与土壤理化因子的关系
如表2所示,各样地土壤孔隙度、土温、总磷和总钾差异不显著;含水量存在显著性差异(F=1.22,P<0.05),刺槐林最高,花椒核桃林次之,苹果林略低于花椒核桃林,耕地最低;电导率存在显著性差异(F=4.33,P<0.05),花椒核桃林最高,为133.75 μS·cm-1,苹果林和刺槐林较接近,分别为128.25和128.00 μS·cm-1,耕地最低,仅为118.00 μS·cm-1;pH在退耕样点和耕地间存在显著性差异(F=3.68,P<0.05),耕地最高,为7.90,其余3个样点较接近;有机质存在显著性差异(F=9.67,P<0.05),刺槐林最高,为2.55 g·kg-1,苹果林和花椒核桃林较接近,分别为0.98和1.21 g·kg-1,耕地最低,仅为0.75 g·kg-1;总氮也存在显著性差异(F=6.39,P<0.05),刺槐林最高,为0.18 g·kg-1,苹果林和花椒核桃林相同,均为0.08 g·kg-1,耕地最低,仅为0.05 g·kg-1.总体来讲,随着退耕还林年限的延长,退耕样点土壤养分的含量显著增加.
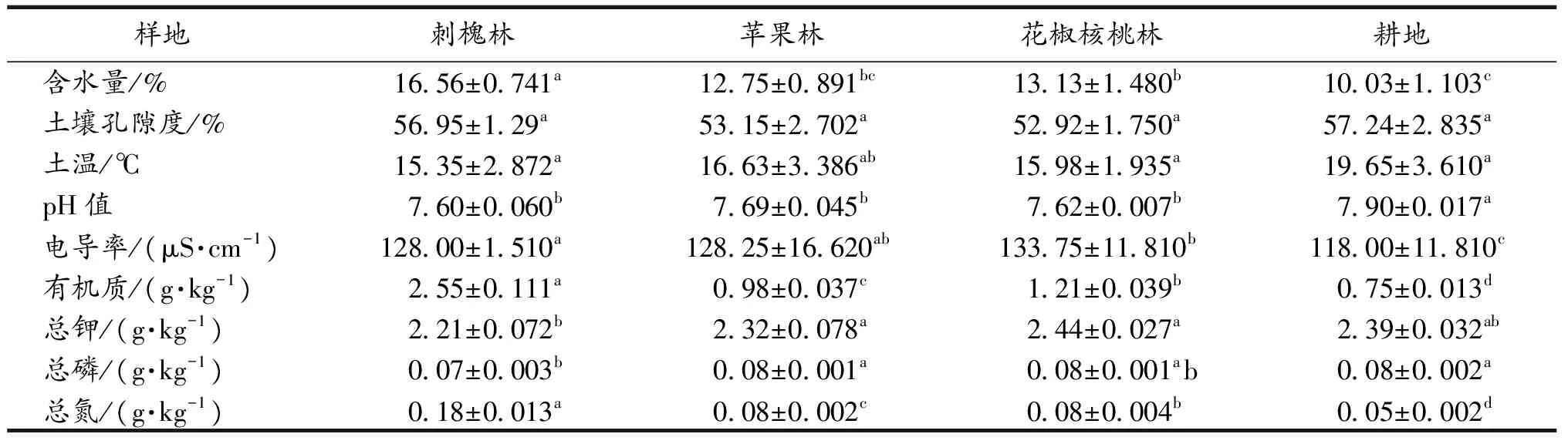
表2 各样点土壤理化因子(平均值±标准误差, n=5)
BIOENV分析结果表明(表3),纤毛虫群落结构受土壤理化因子的综合影响,其中含水量是影响土壤纤毛虫群落结构的主要因子(相关系数r=0.447),其次为含水量、总氮的组合(r=0.442),之后是含水量、有机质的组合(r=0.418),含水量、有机质、总氮的组合相关性也较强(r=0.405).pH、总钾和总磷没有出现在BIOENV分析结果中,说明三者对土壤纤毛虫群落结构的影响不显著.因此,有机质、总氮和含水量是影响纤毛虫群落结构的主要因素.
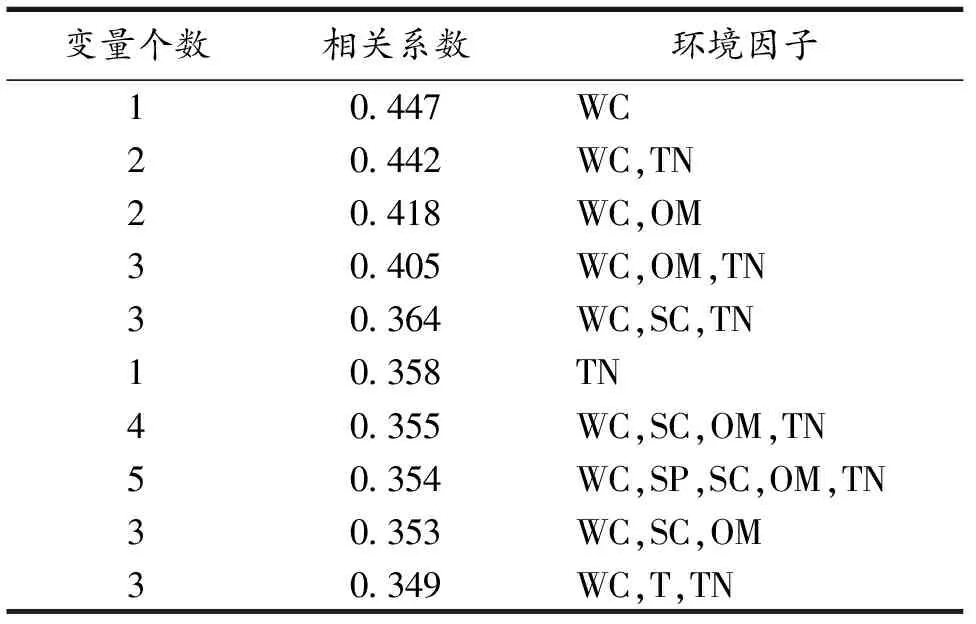
表3 土壤纤毛虫群落与土壤理化因子的 BIOENV相关分析结果
4 讨论
4.1 退耕还林区不同林地土壤纤毛虫群落组成
土壤纤毛虫的群落组成,一方面受地上植被类型的影响,如植物群落组成的复杂程度、腐殖质的组成和质量等是影响土壤动物密度和物种数的重要因素[39].本研究调查的4块样地植被恢复程度不同,土壤纤毛虫的群落组成存在显著差异,退耕样点土壤纤毛虫物种数和平均密度明显大于耕地样点,且刺槐林最优,主要是由于林地植物群落复杂,密度和盖度较大,土壤裸露面积少,水分蒸发量低,相较于耕地,林地更适合土壤纤毛虫生存.赵乌英嘎等[40]的研究发现植物群落能给土壤动物提供小生境,且植被群落复杂程度与小生境及土壤动物的数目呈正比,这与本研究结果相似.另一方面与土壤环境有关,林地的土壤表层积累了大量腐殖质和枯枝落叶,使得土壤动物、微型生物和微生物活动频繁,极大的增加了土壤通气性和孔隙度.除此之外,腐殖质和枯枝落叶分解后产生的有机质等土壤养分对中小型土壤动物个体数具有正向促进作用[26,28,41].退耕还林区的优势类群为高等类群散毛目,而耕地样点的优势类群为低等类群肾形目,这与李涛等[42]的研究结果一致,即与耕地相比,退耕还林所形成的林地可以为土壤纤毛虫提供更理想的生境.耕地中占据主导地位的土壤纤毛虫是较为低等的肾形纲,其能适应恶劣的土壤环境,而退耕样点肾形纲的比例明显下降,高等类群旋毛纲,异毛纲等数量增加,所以耕地的C/P系数显著高于退耕样点,表明高等类群开始占据主导地位,高等类群纤毛虫的生存需要良好的土壤生境.因此,也说明退耕还林后土壤环境得到了显著改善.
4.2 退耕还林区不同林地土壤纤毛虫群落多样性
土壤纤毛虫群落多样性指数是表征纤毛虫群落结构差异的重要指标[43].本研究中,耕地样点纤毛虫Shannon-Wiener指数、Simpson指数、Margalef指数和Pielou指数都显著低于退耕样点(P<0.05),表明研究区退耕还林后林地土壤纤毛虫群落结构趋于稳定,群落组成趋于复杂,多样性明显提高,原因是林地可以给农民带来的经济效益有限,受到的人为因素干扰也相对较少,而耕地人为活动强烈,为了提高农作物产量,人们会采取不同的耕作方式,或间作、轮作等不同的种植方式,这不仅使得地上植被稀疏且单一,还破坏了营养物质含量最高的表层土[44-45],表层土是多数土壤纤毛虫赖以生存的环境.郝宝宝等[46]研究发现退耕还林后林地土壤结构会演化为复杂多样化的生态系统,更加有利于土壤动物生活,进而使土壤动物数量相比耕地有所增加,土壤动物多样性指数与撂荒地有显著性差异,这与本研究结果相似.另外,退耕还林的实施过程中,所选树种的不同会导致植被的恢复和重建产生差异,地下分泌物和地表凋落物为土壤纤毛虫的生存和发展提供了条件[25],不同的植被类型会使得土壤纤毛虫的群落多样性存在差异,刺槐具有固氮的能力,能增强土壤的含氮量,改善耕地土壤环境,这也有利于促进植被生长,加速植被的恢复演替[47],因此刺槐林土壤纤毛虫的群落多样性更高.
4.3 退耕还林区不同林地土壤理化因子对纤毛虫群落的影响
对各样点土壤纤毛虫群落特征参数和土壤理化因子做生物与环境相关分析(BIOENV分析),结果表明,纤毛虫群落结构受土壤理化因子的综合影响,其中,有机质、总氮和含水量是影响纤毛虫群落结构的主要因子.本研究中耕地土壤含水量、有机质含量、总氮等营养物质较另外3个样点低,宁应之等[48]的研究发现土壤含水量是原生动物丰度影响最大的环境限制因子,冯伟松等[49]对南极菲尔德斯半岛地区土壤原生动物的生态学进行了研究,发现土壤原生动物密度与含水量呈极显著正相关,这与本研究结果一致.殷秀琴[50]等的研究也表明,有机质、总氮等土壤养分与土壤动物群落有密切的联系.综上所述,本研究认为相较于耕地,退耕还林后林地中土壤纤毛虫的群落多样性显著提高,且不同林地对土壤纤毛虫群落多样性的影响不同,生态林地上植被以及土壤环境较好,纤毛虫群落多样性最高;经济林受到外界的干扰较多,纤毛虫群落多样性相对较差,但混交经济林略优于单一经济林;耕地人为活动强烈,土壤纤毛虫的群落多样性最差.本研究只对退耕还林年限为17年的林地进行了研究,而退耕还林区土壤纤毛虫的群落特征随退耕还林年限的变化特征还需动态监测.
