基于13C脉冲标记法探究种植密度对侧柏幼苗生长及光合碳分配的影响
2022-05-09孔玉华朱庆征曲安然朱龙飞杨小燕
孔玉华,朱庆征,曲安然,朱龙飞,2,杨小燕
(1.河南农业大学林学院,河南 郑州 450002;2.中建七局建筑装饰工程有限公司,河南 郑州 450000)
森林生态系统作为陆地生态系统最大的碳储存库,在减缓气候变化及固碳放氧等方面具有重要作用。据统计,全球约有46%的陆地有机碳储存在森林生态系统中,其中森林植被有机碳储量占整个陆地植被碳储量的80%~90%[1-2]。植物通过光合作用固定的碳(简称光合碳)是土壤有机碳的主要来源之一,也是大气-植物-土壤系统碳循环的重要组成部分[3-4]。植物通过光合作用将大气中的CO2合成为植物有机碳固定在植物体中,一部分通过植物韧皮部传输到各器官组织用于植物的生长发育,另一部分以根际沉积、根系分泌物等形式输入土壤[5-6]。以往研究表明,光合碳在植物-土壤系统中的分配比例不仅受温度[7]、水分[8]、土壤类型[9]等环境因素的影响,还与植物种类、林分密度[10-11]等生物因素有关。密度制约植物个体对环境资源的吸收利用,是自然界中十分重要的选择压力之一[12]。控制林分密度是人工林管理措施中的一种重要方式,合适的林分密度不仅可以提高木材生产量,改善土壤质量,增加生物多样性,而且有利于人工林生态系统的可持续经营与发展[11-13]。近年来许多学者在林分密度对土壤的碳储量[14]、土壤微生物群落结构[15]、林下植物多样性[16-17]、林木生产力[18]等方面开展了大量研究。然而,林分密度作为影响人工森林生态系统的重要因素之一,其对植物光合碳分配的影响规律仍存在疑问。
13C稳定性同位素标记技术具有标记均匀、安全可靠、无放射性等特点,现已被广泛应用于土壤有机碳的周转、植物光合碳的固定、分配及转移等方面的研究[19-21]。目前,国内利用该技术研究光合碳在植物-土壤系统中的分配多集中在小麦(Triticumaestivum)、玉米(Zeamays)、水稻(Oryzasativa)等农作物方面[22-24],鲜有对林木植物进行标记研究的报道。而定量研究林木树种光合碳在植物-土壤系统中的转移及分配规律,对林业管理和全球碳氮循环研究有重要意义。侧柏(Platycladusorientalis(L.) Franco)因其具有改善土壤结构、增加土壤肥力和良好的固碳能力等生态功能,是我国北方植树造林的主要树种之一[25-26]。徐程扬等[27]研究发现,改变种植密度对侧柏人工林根系产量与垂直分布有极显著的影响。李瑞霞等[28]进一步研究表明,种植密度的变化会改变侧柏人工林根系生物量进而导致生态系统碳储量发生变化。林分密度通过影响植物个体获取的资源数量,进而制约植物的生长与光合碳的分配。因此,本研究拟通过盆栽模拟试验,以树龄1 a的侧柏幼苗为研究对象,利用13C脉冲标记法示踪不同种植密度下侧柏幼苗各组织部位固定的光合碳在植物-土壤系统中的分配比例,分析幼苗各组织部位的生物量和土壤理化性质,以期为林木植物的同位素标记与光合碳分配提供参考依据,并为人工林碳周转机理的研究提供理论依据。
1 材料与方法
1.1 研究区概况
本试验于2019年4月在河南农业大学林业试验站(E 113°42′,N 34°43′)内进行。该区属北温带大陆性季风气候,年均气温15.6 ℃,降水量542.2 mm。8月最热,月均气温25.9 ℃;1月最冷,月均气温2.15 ℃。全年日照时间约1 869.7 h,无霜期209 d。
1.2 试验设计及样品采集
试验材料选取优质的侧柏种子,经1% NaClO浸泡消毒30 min、40 ℃温水浸泡24 h后置于阴凉通风处沙藏15 d,露白后于普通营养钵中进行播种,容器体积22 cm×22 cm×40 cm(上口径×底×高),每盆梅花点6穴,每穴3粒,上覆2 cm基质,轻轻压实,同时6盆不播种作为对照CK。为模拟野外侧柏人工林种植密度,待苗高约5 cm时进行间苗,每盆分别定苗1株、3株、5株,作为低、中、高密度处理,定期浇水和拔草。每个种植密度水平设置8个重复,共计24盆。设置13C脉冲标记处理和未标记处理,两组幼苗摆放距离间隔5 m以上,以避免未标记处理受到污染。栽培基质按草炭:蛭石∶沙子∶壤土=1∶1∶1∶1混合均匀,每盆基质约重15 kg(N∶P∶K=6∶18∶14)。
待幼苗生长1 a后,于2020年6月开始进行同位素脉冲标记,标记期间天气晴朗、光照充足,标记方法参照Lu等[29]。标记过程在特制的有机玻璃箱内进行,每隔10 d通入13CO2气体一次,共标记8次,每次标记时间为上午8∶00~12∶00。标记结束后约30 d进行破坏性取样,先将植株茎叶部用修枝剪完整取出,后将盆中土壤全部倒出,轻抖根茎以收集根际土壤;待根际土壤收集完毕后,用蒸馏水将根系冲洗干净标记装袋后带回实验室。
1.3 样品处理与测定
将带回实验室的植株地上部分与地下部分单独分开,放入105 ℃烘箱内杀青处理20 min后,置于70 ℃烘箱内烘干至恒重,研磨,过0.15 mm筛备用。将采集的根际土壤过2 mm筛后,分为2份。1份置于4 ℃冰箱保存,用于后续土壤活性碳氮的测定;另1份置于室内风干至恒重用于土壤pH、颗粒组成体积分数的测定,后过0.15 mm筛用于13C、全碳(TC)、全氮(TN)等的测定。
采用同位素质谱仪(Finnigan MAT253,Thermo Electron)测定土壤和植物样品的δ13C 值;采用EPSON PERFECTION V700 Photo根系扫描仪对侧柏根系进行扫描,并结合WINRHIZO PRO 2007根系分析系统(专业版)统计根系数据;土壤pH采用10 g风干土与25 mL蒸馏水(土∶水=1∶2.5)混合摇匀后用酸度计测定;土壤颗粒组成体积分数(黏粒<0.002 mm;粉粒0.002~0.05 mm;砂粒0.05~2 mm)采用土壤粒径粒型测量系统(Mastersizer 2000,Malvern,英国)测定;土壤TC、TN含量采用元素分析仪(Euro Vector EA3000,意大利)测定;用1 mol/L KCl溶液浸提新鲜土壤(土∶液=1∶5,w/w),采用SKALAR流动分析仪(SAN++,荷兰)分析浸提液土壤溶解性有机碳(DOC)、溶解性有机氮(DON)、铵态氮(NH4+-N)和硝态氮(NO3--N)含量。
1.4 数据处理与分析
自然土壤或植物的自然丰度用δ13C值来表示,计算公式如下:
δ13C(‰)=(Rsample/RPDB-1)×1000
式中:Rsample代表13C/12Csample,RPDB代表13C/12CPDB,为0.011 237 2。
标记后的土壤或植物一般采用Atom(%)来表示,Atom(%)与δ13C(‰)相互转换的公式如下:
Atom13C(%)= [(δ13C+1000)×RPDB] / [(δ13C+1000)×RPDB+1000]×100%
侧柏幼苗各组织部位固定13C量计算公式如下:
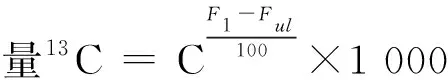
式中:Fl为标记组织部位13C丰度,Ful为未标记组织部位13C丰度。
数据采用SPSS 19.0(IBM,纽约,美国)软件对不同处理进行差异显著性检验(Duncan),并对侧柏幼苗13C含量、根系形态和其他土壤指标进行相关性分析,用Excel 2013软件制图。
2 结果与分析
2.1 种植密度对侧柏幼苗生长的影响
由表1可知,种植密度显著影响了侧柏幼苗根系、根尖数、根系表面积和根总长(P<0.05)。随着种植密度的增加,根系的这些指标呈递增的趋势,但未呈倍数增加。随种植密度的增加,地上部分生物量、地下部分生物量和总生物量均呈递增的趋势,变化范围分别为26.6~39.3、17.3~26.2和43.8~65.5 g/盆。高密度和中密度的侧柏幼苗地上部分及总生物量显著高于低密度(P<0.05),但其二者之间无显著差异。从低密度到中密度,地上部分生物量的增幅最大,增长率为42%,而中密度到高密度增幅仅为29%。高密度种植侧柏幼苗地下部分生物量显著高于低密度处理(P<0.05)。

表1 不同种植密度下侧柏幼苗根系形态与生物量
2.2 种植密度对植物-土壤系统碳氮含量及土壤理化性质的影响
分析表2可见,相较于未脉冲标记处理,13C脉冲标记后所有种植密度下侧柏幼苗地上和地下部分13C含量均呈较显著富集,地上部分分配的13C含量显著高于地下部分和土壤(P<0.05)。本次标记使侧柏幼苗地上和地下部分的δ13C值达到了+100.0‰ PDB以上,表明通过本次13C脉冲标记1 a侧柏幼苗可采用较低的用量,为后期土壤碳组分周转实验提供科学材料。随种植密度的提高,侧柏幼苗单位质量地上部分、地下部分和土壤中的13C含量略有下降,但并未达到显著性差异。植物-土壤系统各组织部位的TC、TN含量均呈地上部分显著高于地下部分和土壤(P<0.05),C/N则呈地下部分>地上部分>土壤的趋势(P<0.05)。种植密度对各组织部位的TC、TN含量和碳氮比(C/N)有一定影响,但并不显著。

表2 不同种植密度下侧柏-土壤系统各组织部位碳氮含量
植物根系通过根际沉积等形式将根部的有机、无机化合物释放到周围土壤中进而影响土壤的理化性质。由表3可知,与原土壤(CK)相比,侧柏的种植显著增加了土壤中NH4+-N 、DOC、DON含量,降低了土壤NO3--N含量(P<0.05),对土壤pH和土壤粒径体积分数无显著影响。高密度种植显著提高了土壤DOC、DON含量,降低了土壤黏粒体积分数(P<0.05)。

表3 不同种植密度对土壤理化性质的影响
2.3 种植密度对13C含量在侧柏-土壤系统中分配比例的影响
由图1可知,各种植密度下的侧柏幼苗地上部分、地下部分和土壤中13C含量变化范围分别为:953.51~980.32、585.61~526.87 和0.02~0.27 mg/盆。随种植密度的增加,在植物-土壤系统中13C总量呈降低趋势,低密度处理的13C总量最大,为1 539.14 mg/盆,高密度的13C总量最小,为1 502.18 mg/盆,但不同处理间无显著差异。种植密度显著影响了13C含量在植物-土壤系统中的分配比例,其中植物地上部分所占比例最高。随着种植密度增加,地上部分13C含量的分配比例呈增加的趋势,变化范围为61.95%~64.92%;地下部分则呈降低的趋势,分配比例为38.04%~35.07%;根际土壤中13C含量最低且无显著变化,仅占0.01%~0.02%。

图1 不同种植密度下13C含量在植物-土壤系统中的分配比例Figure 1 The distribution ratio of 13C content in plant-soil system under different planting densities
2.4 侧柏幼苗13C含量、根系形态与土壤因子间的相关性分析
为进一步研究侧柏幼苗光合碳分配的影响因素,对各指标进行了相关性分析。由表4可知,侧柏幼苗地上部分13C含量与地下部分13C含量呈极显著正相关(P<0.01),与土壤中13C含量呈显著正相关(P<0.05)。植株地上与地下部分的13C含量均与土壤粉粒体积分数呈显著正相关、与砂粒体积分数呈显著负相关性关系(P<0.05)。侧柏植株地上与地下部分及总生物量与侧柏幼苗的根尖数、根系表面积、根体积、根总长均表现为极显著正相关关系(P<0.01)。
3 讨论
植物根际碳沉积是土壤有机碳的重要来源之一,植物通过光合作用将光合产物碳以根际沉积的方式输入土壤,进而影响土壤碳储量及其理化性质[30]。本研究中,高种植密度显著增加了侧柏幼苗地上及地下部分生物量(表1)和土壤DOC含量(表3),表明侧柏幼苗高密度的种植显著增加根际碳输入,这与王艳芳等[31]研究结果一致,暗示地下部分生物量的增加影响根际微生物群落的结构和活性,还增加了植物根系脱落物的碳输入量,从而有利于土壤的固碳效应[32]。胡凯等[33]研究也表明植物根系生物量与根际碳输入、微生物群落的组成和数量呈正相关关系,再次验证了本研究的推测。氮素是植物生长发育所必需的营养元素之一。本研究中随着种植密度增加,土壤NO3--N和DON含量均呈显著增加的趋势(表3),这可能是由于地下生物量的增加提高了植物根系分泌物含量,进而为根际土壤提供了更多的氮源[34]。其次,中密度和高密度处理下土壤C/N较低(表2),有利于土壤中有机质矿化作用的进行,提高了土壤中活性氮含量[35]。
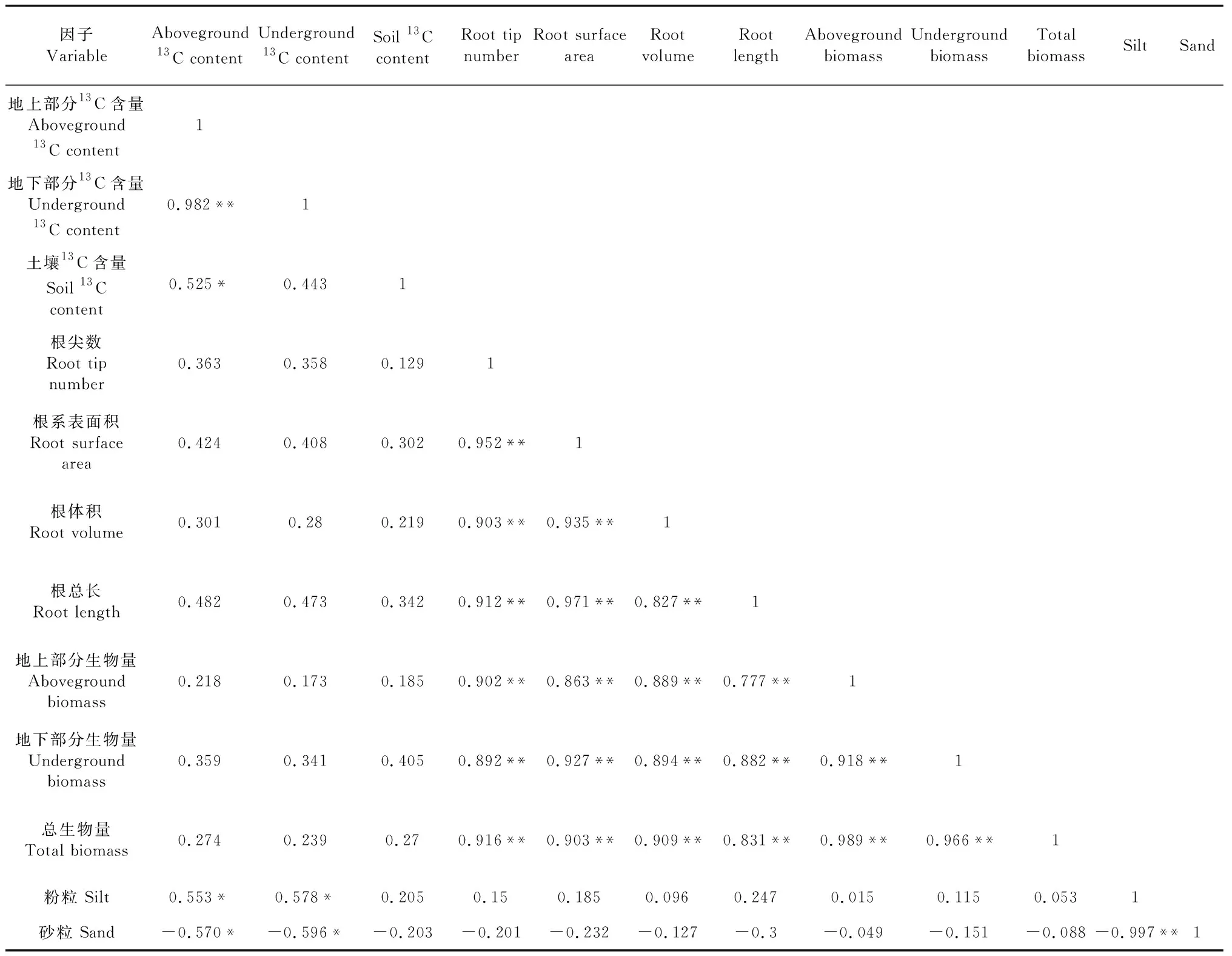
表4 侧柏幼苗13C含量、根系形态与土壤因子间的Pearson相关系数(n=9)
13C稳定性同位素标记技术目前已被广泛应用于生育期、作物品种以及施氮量等条件下植物光合产物分配的研究。本研究中,经过8次13C脉冲标记后,侧柏幼苗地上与地下部分的δ13C值均达到+100‰以上,分配比例呈地上部分>地下部分>土壤的趋势(图1),这与于晓燕等[36]的研究结果一致。Lu等[29]对水稻进行13C脉冲标记后,也发现其光合碳主要存留于地上部,小部分分布于根系中。随着种植密度增加,侧柏幼苗地上及地下部分13C含量呈下降的趋势(表2),这主要是由于中密度和高密度水平下幼苗的总生物量显著高于低密度水平(图1),而生物量的增加相当于对13C含量起到稀释作用[37]。安婷婷等[37]对玉米进行13C标记1 d后,测得其茎叶和根中13C含量为1 568‰和598‰,与本试验结果相差10倍。而尹云锋等[20]对水稻进行5次标记后发现其茎叶和根中13C含量为1 377‰和1 413‰。于晓燕等[36]对树龄1 a杉木幼苗进行4次标记后发现其针叶、枝干和根中含量为116.74‰、116.8‰和124.08‰,与本试验结果相似。以上研究表明植物对13C富集程度的差异可能与标记次数、标记时间和植物种类等因素有关。与玉米、水稻等禾本科植物相比,树龄1 a的侧柏幼苗的生育期长、茎叶和根生物量小,因此其光合碳富集程度远低于水稻和玉米。
侧柏植株通过叶片光合作用将13CO2吸收固定在植物体内,后在植物-土壤系统中进行转化和再分配。本研究中侧柏幼苗经脉冲标记后,其地上部分13C含量与地下部分及土壤中13C含量呈显著正相关(表4),表明光合碳在植物-土壤系统中的分配与流动具有一定的正关联。种植密度的提高显著增加了光合碳在地上部分的分配比例,降低了地下部分的分配比例,而土壤中的比例无显著差异(图1),这可能是因为低密度种植水平下侧柏幼苗所需养分和能量较少,在满足地上部分生长所需后,侧柏幼苗将其余固定的13C转运至根部;而中密度和高密度种植水平下侧柏幼苗地上部分生物量显著增加,相应所需利用的13C含量较多,从而在植物个体间产生了种内竞争作用,为了植物个体更好的生长,便增加了地上部分光合碳的分配比例,而降低了向根部的转运。Kuzyakov等[10]研究表明玉米光合碳有0.5%~10%被转运到土壤中,邓扬悟等[11]研究发现水稻土壤中光合碳比例仅占3.8%~10%。本试验条件下,侧柏土壤中13C含量较低,仅占0.01%~0.02%,一方面可能是因为1 a侧柏仍处于生长发育期,被同化的13C大部分用于植物的生长,极小部分通过植物根系分泌至土壤中,另一方面可能是由于本试验未考虑土壤和侧柏根系的呼吸作用所致[37]。
4 结论
本试验条件下,高种植密度水平显著提高了侧柏幼苗的地上部分、地下部分与总生物量,土壤DOC、DON 和NO3--N含量(P<0.05)。侧柏植株地上与地下部分及总生物量与侧柏幼苗的根尖数、根系表面积、根体积、根总长均呈极显著正相关关系(P<0.01),且侧柏幼苗地上部分13C含量与地下部分13C含量及土壤13C含量呈显著正相关关系(P<0.05)。种植密度显著影响13C含量在各组分的分配比例,随种植密度的提高,分配在地上部分的比例呈递增趋势,分配在地下部分的比例呈递减趋势,而分配在土壤中的比例无显著差异。
