大白菜CMS不育类型的鉴定及细胞学观察
2022-05-05王晓玲李庆飞原让花王梦梦肖艳董彦琪常怀成张倩倩
王晓玲 李庆飞 原让花 王梦梦 肖艳 董彦琪 常怀成 张倩倩




摘要:探求大白菜 CMS 不育类型及发生的时期、特点,利用 PCR 扩增方法鉴定 CMS 的类型,并采用石蜡切片的方法观察花蕾不同长度时花药的发育情况。结果表明,I 晋毛不育系、II 苔不育系为 Polima 型雄性不育系类型,III紫不育系为 Ogura 型雄性不育系类型;保持系花粉皆正常发育,I 晋毛不育系花蕾在长度小于等于 1 mm 时就明显观察到花药发育异常,II 苔不育系和 III 紫不育系可以形成花粉囊但大多呈现畸形,在后期花药内出现多药隔、无药隔或花粉粘连在一起使其无法进行正常散粉。综上,3 种 CMS 不育系皆不属于 nap 型雄性不育系;花蕾中花粉败育的时期很早,当 L≤1 mm 时就决定了花药能否形成花粉囊产生花药。本研究为利用 CMS 不育系培育新品种奠定了基础。
关键词:大白菜;CMS;类型鉴定;石蜡切片;花粉;细胞学观察
中图分类号:S634.1
文献标志码:A
文章编号:1673-2871(2022)03-021-05
Abstract:In order to exam the sterility type, sterility onsetting time and sterility characteristics of CMS Chinese cabbagelines, PCR amplification and paraffin section of flower buds were conducted in this experiment. The results showed thatJinmao I male sterile line and Tai II male sterile lines were Polima type male sterile, and Purple III male sterile line wasOgura type male sterile. The pollen of maintainer lines developed normally. Abnormal anther development was observedwhen the flower bud of Jinmao male sterile line reached less than or equal to 1mm. Pollen sacs were formed in Tai II malesterile line and Purple III male sterile line, but most of them showed deformity. In the later stage the pollen sac had multi ple septa, no septum or pollen adhered together, so that pollen grains could not disperse normally. These results suggestthat three CMS male sterile lines evaluated in this experiment do not belong to nap type male sterile. The pollen abortioninitiated at very early stage of flower development, flower bud less than or equal to 1mm. Our discoveries provided afoundation for new Chinese cabbage CMS male sterile line development.
Key words:Chinese cabbages; Cytoplasmic male sterility; Type identification; Paraffin section; Anthers; Cytologicalobservation.
在十字花科作物大白菜的杂交优势育种中,利用其雄性不育系杂交育种是一条非常重要的途径[1-2]。雄性不育性在高等植物中普遍存在,雄性不育可以分为细胞质不育型、细胞核不育型和核质互作不育型3种类型[3]。由于大白菜资源中没有细胞质雄性不育源,生产中使用的细胞质雄性不育系均来源于异源细胞质[4]。细胞质雄性不育系主要有3种类型:萝卜 Ogura CMS、甘蓝型油菜 Polima CMS和甘蓝型油菜nap CMS[5]。雄性不育系的花粉由于受多种因素的影响,花粉不能正常的发育或者没有生活力,雄蕊的败育会因品种或类型的不同而有所差异[6],不同大白菜材料的CMS和保持系的线粒体DNA的RFLP、RAPD和AFLP呈多态性,CMS和保持系的线粒体基因组结构也存在着显著差异[7]。产生细胞质雄性不育系的分子机制主要有两种:一是线粒体 DNA 产生新的阅读框(open reading frag-ment,ORF),产生有毒的蛋白,从而影响到线粒体的正常功能;二是与线粒体雄性不育有关的基因与正常线粒体基因重组或共表达,直接或间接影响正常基因的功能,产生雄性不育。如在Ogura型萝卜中发现的orf138、nap甘蓝型油菜中发现的orf222、Po lima 型油菜中发现的 orf224、叶用芥菜中发现的orf220等都是在不育系材料中发现的与不育性相关的开放阅读框。
细胞和组织在离开机体后很快会发生溶解和腐败,很难维持它原有的形态和结构[8-9],因此,通过固定、脱水、透明、浸蜡、包埋、切片、脱蜡、染色等步骤制作石蜡切片可免使细胞组织死去,以便于后期进行观察分析[10]。
本试验在以往研究的基础上,探求大白菜细胞质雄性不育发生的时期及其特点,对现有的不育系材料进行鉴定分类和细胞学观察。采用同源序列基因扩增技术,依据与细胞质雄性不育有关的线粒体嵌合基因orf224、orf138、orf222设计出3对引物, 对3种大白菜花蕾粒线粒体DNA进行PCR扩增,从而获得差异片段,并通过电泳试验对大白菜细胞质雄性不育系的类型进行鉴定;继而采用石蜡切片的方法,用光学显微镜观察花蕾在不同長度时花药的发育程度。为细胞质雄性不育系在大白菜新品种选育中的应用奠定理论基础。
1 材料与方法
1.1 材料
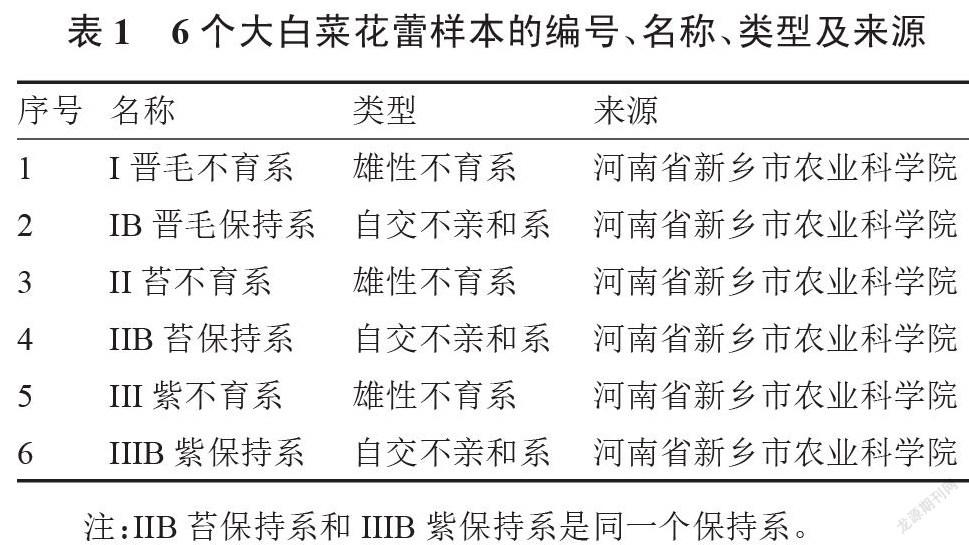
本试验采用3个不育系及相对应的保持系(表1)。
1.2 器材与试剂
器材:光学显微镜、真空抽气机、恒温箱、蜡片切片机、相机、温台、展片台、蜡杯、酒精灯、载玻片、盖玻片、毛笔。
试剂:FAA固定液、乙醇溶液、醋酸、二甲苯、甲醛、伊红、粘片剂、石蜡、染色液(固绿、番红)、加拿大树胶。
1.3 方法
于2019 年 12 月 10 日进行冬季育苗,2020 年 2 月25日定植于大棚,每份材料种植40株,2020年4 月20日在新乡市农业科学院研发中心试验大棚进行采样,并在此研发中心实验室进行室内试验。采样前注意先将塑料袋和镊子在液氮中预冷,用镊子将大白菜花蕾粒轻轻采下放入采样袋中。根据试验田种植的材料,取配对的3个不育源,3个与其对应的保持系。在种植的植株上,完全随机选择花序,每个材料取顶部花蕾3~6个,取样完毕后将样品放入液氮罐中,于超低温冰箱-80 ℃冷冻储存。不育系类型的鉴定和细胞学观察于河南科技学院园艺园林学院分子实验室进行,采用混合样,无重复。
1.3.1 大白菜细胞质雄性不育系类型的鉴定 在NCBI中检索的细胞质雄性不育有关的线粒体嵌合基因 orf224(DQ400846.1 )、orf138、orf222,利用生物学软件primer5软件设计出3对引物,由上海生物技术生物工程有限公司进行合成。使用改进的CTAB方法[11]分别提取6个大白菜花蕾粒样本花蕾DNA;并对其进行PCR扩增,扩增产物经2.0%琼脂糖凝胶电泳检测,鉴定其细胞质雄性不育系的不育类型。
1.3.2 大白菜细胞质雄性不育系的细胞学观察 于2020 年 5 月 6 日 9:00—10:00 采取长度分别为(L≤ 1 mm、1 mmm<L≤2 mm、2 mm<L≤3 mm,L代表花蕾的长度)新鲜大白菜花蕾,此时植物细胞分裂活动明显,取材效果最好。为了维持大白菜细胞的自然状态,使形态和结构不发生变化,取材后参照段国禄等的固定方法马上进行固定,参考何艳艳、张宝华 的 方 法 制 作 石 蜡 切 片 [12- 14]。 在 光 学 显 微 镜(XSP-8C,上海精密仪器仪表有限公司)下观察切片,并进行拍照记录。
2 结果与分析
2.1 引物设计结果
从 NCBI 获得线粒体嵌合基因 orf138、orf224、orf222的序列,并通过Primer 5软件设计引物(表2)。
2.2 CMS类型的鉴定
用设计的 orf138 引物、orf224 引物和 orf222 引物对3个大白菜细胞质雄性不育系及其保持系的6份花蕾粒DNA进行扩增。结果如图1所示,orf138引物扩增出的条带2清晰可见,即Ⅲ(紫不育系)为Ogura 型雄性不育系类型,ⅢB 为紫保持系;orf224引物扩增出的条带4和条带6很清晰,即Ⅰ(晋毛不育系)、Ⅱ(苔不育系)为Polima型雄性不育系类型;条带5(ⅡB)、条带7(ⅠB)分别为苔保持系、晋毛保持系。orf222引物未扩增出条带,说明Ⅰ(晋毛不育系)、Ⅱ(苔不育系)、Ⅲ(紫不育系)都不属于nap型雄性不育系类型;orf224、orf138和orf222对保持系材料扩增条带均不清晰,即基因序列中没有与细胞质雄性不育相关的开放阅读框序列。试验结果表明:Ⅰ(晋毛不育系)、Ⅱ(苔不育系)为Polima型雄性不育系类型,Ⅲ(紫不育系)为Ogura型雄性不育系类型;Ⅰ(晋毛不育系)、Ⅱ(苔不育系)、Ⅲ(紫不育系)都不属于nap型雄性不育系类型。
2.3 保持系材料和不育系材料花药发育的特点
大白菜正常的花药由4个药室组成,药室由外到内分别是表皮、纤维层、中层、绒毡层。绒毡层呈长方形规则紧密地排列在药室最里层,主要为花粉母细胞提供营养,随着花粉发育会逐渐降解,花药同一侧 2 个相邻小孢子囊之间的组织也会逐渐消失,打通变成1个药室。通过切片观察,可以看到保持系材料 IB 晋毛保持系、IIB 苔保持系和 IIIB 紫保持系的花药在各个时期都发育正常(图2~4)。
大白菜CMS不育主要是雄蕊发育不正常,不能产生可育的花粉。通过切片观察可以看到,不育系材料中大多在花蕾长度≤1 mm时就可以明显看到差异,而后期本该形成药室的地方只存在大量薄壁细胞。个别药室可以形成花粉囊产生花粉,但在后期因花粉延迟发育或粘连在一起无法正常散粉。此外,还出现个别花药有多药隔、无药隔情况(图2~4)。
2.4 保持系材料和不育系材料花药发育的区别
根据对保持系材料与不育材料花药切片的细胞学观察,在花蕾长度小于等于1 mm时,保持系材料 IB 晋毛保持系(图 2-2)、IIB 苔保持系(图 3-2)、IIIB紫保持系(图4-2)在花蕾的横切面上,可见1个雌蕊和4个雄蕊,花药呈蝶形,4个药室已经形成,可以看见绒粘层在花粉囊内染色较深,且维管束和花粉囊内细胞均被染色。而I晋毛不育系(图2-1)和II苔不育系(图3-1)的4个花粉室尚未形成,III紫不育系(图4-1)可以看见有花粉囊的形成,但染色较浅,没有产生明显的花药。
当花蕾长度为 1 mm<L≤2 mm时,保持系材料花药中药室、药隔更加清晰,绒粘层整齐的排列在小孢子囊母细胞的周围,可以看见清楚的药室由外到内分别为表皮、药室的内壁、中层、绒毡层(图 2-4,3-4,4-4)。但不育系I晋毛不育系(图2-3)和II苔不育系(图 3-3)在这个时期仍然没有形成花粉囊,III紫不育系虽然有花粉囊的形成,但花粉囊内的花粉较少,个别花药内不能正常形成4个药室,还有个别花粉囊内绒毡层膨大而挤压小孢子形成一条缝隙,造成小孢子发育不良,呈现败育的现象(图 4-3)。当花蕾长度为2 mm <L≤3 mm时,保持系材料孢间层逐渐溶解,细胞逐渐分开,变成圆球形,个别细胞呈四边体形,着色较深(图2-6,3-6,4-6)。不育系材料I晋毛不育系雌蕊发育正常,从此时雌蕊发育的程度以及IB晋毛保持系的发育程度来看,花粉发育应该和它的保持系发育一样,但此时I晋毛不育系仍然没有药室形成,细胞呈半透明状,是典型的无花粉囊(图2-5)。II苔不育系发育较晚,只有角隅处可以形成2~3个药室,而且花蕾横切面不能呈正常的蝶形(图3-5)。而可以观察到III紫不育系花粉囊内的细胞染色較浅,花粉囊内的花粉粘连无法形成单个的圆形花粉粒,个别出现无药隔的现象(图4-5)。
3 讨论与结论
近20年来,在大白菜细胞质雄性不育性研究方面,已报道有 Ogura CMS、Polima CMS 和 nap CMS等多种细胞质雄性不育系类型。Polima CMS是最有生产价值的细胞质雄性不育系,不仅在大白菜CMS不育系的选育中被广泛利用,而且在其他十字花科作物雄性不育系的选育中也被广泛应用[15]。2005 年赵利民[16]建立了一套大白菜新品种灰色关联度多维综合评判方法,为客观、准确地从众多大白菜组合中筛选出优良新品种提供了科学依据,提高了新品种选育的准确性和选择效率。2018 年崔冰等[18]对大白菜细胞质雄性不育形成的机制进行研究。2020 年杨红丽等[19]培育出耐热抗根肿病萝卜胞质雄性不育多基因聚合新品种。随着科学技术的发展,准确快速鉴定样本所归属的雄性不育系的类型,以便为开展细胞质雄性不育系在育种中的应用奠定基础,并为实现大白菜优势杂交育种提供分子依据。笔者对 6 个大白菜材料 CMS 的不育类型进行鉴定,发现其中Ⅰ晋毛不育系、Ⅱ苔不育系为Polima 型雄性不育系类型;Ⅲ紫不育系为 Ogura 型雄性不育系类型;因其基因序列中没有与本试验CMS 不育相关的开放阅读框序列 ,所以 orf224、orf138和orf222引物均未扩增出清晰条带。
通过石蜡切片法进行观察发现本试验中大白菜保持系花药均发育正常,可以形成4个蝶形药室从而产生可育的花粉。I晋毛不育系与可育材料IB晋毛保持系比较,I 晋毛不育系的败育花药一直处于孢原细胞之前,在整个时期内都没有形成花粉囊,属于典型的无花粉囊型败育,它的药室异常出现较早,当花蕾长度为L≤1 mm时差异显著;II苔不育系药室异常出现较晚,在花蕾长度为 1~2 mm 时还未形成花粉囊,发育较晚,在花蕾长度为2~3 mm时只有2~3个花粉囊可以形成,且花药畸形,无法形成正常的蝶形药室,个别花药内还出现多药隔现象;III紫不育系,在后期可以明显地观察到不育系花粉囊内的花粉因相互粘连无法形成单个的花粉粒。2016 年郭瑛琪等[19]大白菜核不育花药败育起始于减数分裂期绒毡层结构异常,小孢子在四分体时期开始败育;不育发生后,伴随着绒毡层细胞异常膨大,严重空泡化,提前降解,小孢子变形干瘪而死亡。2012 年许明等[20]研究表明不育花蕾长度在2.5 mm后就已经败育,败育时期可能发生在减数分裂后期到四分体时期。本试验与其研究结论一致,花蕾中花粉败育的时期较早,当L≤1 mm时,它就决定了花药能否形成花粉囊从而产生花药。
参考文献
[1] 孙昊.橘红心大白菜细胞质不育系的选育与评价[D].沈阳:沈阳农业大学,2018.
[2] 闫世江,张继宁,刘洁.雄性不育在蔬菜杂交育种上的应用[J].蔬菜,2011(10):44-47.
[3] 袁鹤.大白菜雄性核不育的细胞学观察及基因定位和 AFLP分子标记[D].河北保定:河北农业大学,2009.
[4] 张文翔.白菜薹细胞核雄性不育基因的定位[D].沈阳:沈阳农业大学,2019.
[5] 陈烨丽,陈学好,陈锦秀,等.甘蓝细胞质雄性不育类型的分子鉴定[J].上海农业学报,2009,25(3):19-22.
[6] 龙欢,姚家玲,涂金星.3 种甘蓝型油菜雄性不育系花药发育的细胞学研究[J].华中农业大学学报,2005,24(6):570- 575.
[7] 李泽福,夏加发,唐光勇.植物雄性不育类型及其遗传机制的研究进展[J].安徽农业科学,2000,28(6):742-746.
[8] 陈土娟,黄子娟,潘海玉,等.茴香成熟叶柄石蜡切片技术的研究[J].韶关学院学报,2014,35(12):41-45.
[9] 程圆.三种不育源乌菜转育后代生理生化及其花药细胞形态学研究[D].合肥:安徽农业大学,2016.
[10] 王金平,张俊梅.白头翁花的石蜡切片制作[J].信阳师范学院学报(自然科学版),2008,21(4):573-576.
[11] 李荣华,夏岩石,刘顺枝,等.改进的 CTAB 提取植物 DNA 方 法[J].实验室研究与探索,2009,28(9):14-16.
[12] 段国禄,施江.植物制片、标本制作和植物鉴定[M].北京:气象出版社.2008.
[13] 何艳艳.石蜡制片的方法、技巧与注意事项[J].农业研究与应用,2014(4):54-57.
[14] 张宝华.聚焦植物石蜡切片制作[J].吉林省教育学院学报,2013,29(4):153-154.
[15] 董美云.大白菜溫敏雄性不育的温敏特性及选育研究[D].陕西杨凌:西北农林科技大学,2008.
[16] 赵利民.大白菜胞质雄性不育系选育、利用及采种技术的研究[D].陕西杨凌:西北农林科技大学,2005.
[17] 崔冰,石绍峻,李鑫,等.大白菜细胞质雄性不育研究进展[J].山东农业科学,2018,50(3):154-157.
[18] 杨红丽,徐学忠,胡靖锋,等.耐热抗根肿病萝卜胞质雄性不育多基因聚合新品种 CHR18003 的选育[J].陕西农业科学,2020,66(7):67-70.
[19] 郭瑛琪,崔海芳,李海燕,等.大白菜细胞质雄性不育系选育及其细胞学观察[J].云南农业大学学报(自然科学),2016,31(3):427-432.
[20] 许明,郑鹏婧,张欣,等.大白菜细胞核雄性不育甲型“两用系”细胞学观察[J].西北农业学报,2012,21(4):94-98.
