黄精基源植物种质多样性及其利用研究进展
2022-04-20尹舒雅钱丽华
尹舒雅,钱丽华
(杭州市农业科学研究院,浙江 杭州 310024)
黄精(药物名:Polygonati Rhizoma)别名鸡头参、黄鸡、白豆子,是中国特有的传统中草药,始载于晋代《名医别录》,中医理论认为其味甘性平,归脾、肺、肾经,具有补气养阴、健脾、润肺、益肾的功效,是数十种滋补性中医复方或中成药的重要组分[1]。现代科学研究证明,黄精的根茎含有多种活性成分,如黄精多糖、黄精多酚、生物碱、黄酮和甾体皂苷等[2-3],具有抑菌抗炎、调节免疫力、抑制肿瘤、抗氧化、清除自由基、降血糖、调血脂、改善失眠抑郁等多种作用[4,8]。作为药食同源中药,黄精的肉质根状茎肥厚,含有大量淀粉、糖分、脂肪、胡萝卜素、蛋白质、维生素和多种其他营养成分,具有保健功能,久服可使人气力倍增、肌肉坚实、壮骨强筋,对体能恢复十分有益[15]。
根据《中华人民共和国药典》规定,中药材黄精来源于黄精(Polygonatum sibiricum Red.)、滇黄精(P.kingianum Coil.et Hemsl.)和多花黄精(P.cyrtonema Hua.)这3个物种的干燥根茎[1]。近年来随着黄精药用价值和营养保健等功能不断被发掘,黄精原料的需求量日益增加,但黄精野生资源长期处于私挖滥采的状态,野生资源逐步枯竭,虽已有一定规模的人工栽培,但仍处于种源混杂,管理粗放的状态,存在大量依赖采挖的野生黄精作为栽培种苗等问题,有待进一步发展[16]。为更好地保护并开发利用好这一珍贵药材资源,本文主要总结了黄精基源植物种质的多样性及在基因功能、品种选育等方面的研究进展,以期为黄精的栽培和品种改良提供参考。
1 黄精基源植物种质的多样性
1.1 黄精基源植物种质生态分布与表型多样性
药材黄精的3 种基源植物都属于黄精属,黄精属(Polygonatum)传统上属于百合科(Liliaceae),但近年来分子生物学证据支持应将其归入天门冬科(Asparagaceae)[17]。黄精属植物迄今已发现60 余种,分布于中国境内的共有39种,分布于东三省、河北、河南、山西、内蒙古、陕西、宁夏、甘肃、山东、安徽和浙江等省,生长于海拔200~2 000 米的林下、灌丛、水源边、荒石等阴湿环境中[18]。3 种黄精属植物的分布范围、形态特征和生活习性各有不同。根据中国植物志等记载,黄精和多花黄精分布于低海拔山区,其中多花黄精分布于中南地区及相对温暖的江苏、安徽、浙江、江西、福建等地,叶互生,根状茎常结节成块,形似生姜,故习称山姜或姜形黄精;黄精则分布较为广泛,除东北、华北地区外,陕西、宁夏、甘肃等地也有分布,地上部分叶轮生,地下部根状茎一头粗一头细,状似鸡头,因此也俗称鸡头黄精;滇黄精的分布范围较窄,自然产区为云南、四川、贵州等西南部省份,海拔1 400~2 100 米的亚热带地区,其尖端拳卷,且具有黄精属中较为少见的粉红色花被及红色浆果,其根茎结节明显,多连珠状,故又称节节高[18,21]。
近年来,随着对各地种植资源的收集,发现同一物种不同种源的黄精表型存在巨大差异。彭星星等[22]野外调查中发现,多花黄精存在至少5 种形态的花,叶片则根据是否有蜡质,背面是否有毛,背面是否略带紫色以及叶片长宽比呈连续变化,难以清晰分类,根状茎则至少存在细圆柱状、大圆柱状、圆锥状、细瘦连珠状、肥厚连珠状、不规则块状6 种形态,这种变化与经纬度有一定相关性,但同一区域内多花黄精根状茎形态也存在分化,这种分化影响了根状茎有效成分含量,结节状不明显的根状茎总多糖含量显著高于结节状明显的根状茎。刘校分析了来自14个不同产地的黄精植株,发现黄精的地上部随环境变化较为规律,黄精的株高与海拔高度呈正相关,叶片的宽度与海拔呈负相关,开花轮数与海拔呈正相关,而每一轮的花序开花数与海拔无明显特异。不同地区黄精根状茎转换段直径、伸展段形态、根状茎长宽比都有不同,在其分布区域内,根状茎变得肥厚,长宽比缩短,转换段由东南向西北方向逐渐增大,由长鸡头形而逐渐缩短节间变为短鸡头形,采集自湖北省十堰市房县野人谷的黄精根状茎甚至因为伸展段较短呈现出连珠状,近似多花黄精,同时根状茎颜色会由浅白逐渐加深变为黄色,含水量减少,折干率增加[23]。说明除遗传因素外,环境条件对黄精基源植物表型形态特征有着较为显著的影响,因此栽培条件的研究尤为重要。根据对3种黄精基源植物及黄精属内其他物种的表型观察,黄精属植物随地理气候区域变化呈现较为连续的变化,最适生境不同的黄精属物种分布范围间重叠较多,推测黄精属内物种应正处于物种分化过程中[22-23]。
1.2 黄精基源植物种质农艺性状的多样性
3 种黄精基源植物的栽培条件和农艺性状也存在一定差异。黄精为中偏阳性草本植物,多花黄精为中性草本植物,有很好的耐阴性,在大棚、林下等环境中净光合速率最高,黄精其次,滇黄精最低,但在高温强光的全日照环境中,多花黄精净光合速率表现出叶片发黄早衰,净光合速率低于黄精,黄精则生长旺盛[24,26],这为黄精3 种基源植物的栽培模式提供了参考,如多花黄精可栽培于林下,也可与瓜蒌、葡萄、猕猴桃等套种。罗春梅[27]等对比了黄精、多花黄精、滇黄精的物候期,发现滇黄精出苗最早,多花黄精最晚,但仅相差6 天,开花时间最早是黄精,最晚是滇黄精,相差25 天;结实最早是黄精,最晚是滇黄精,相差18天;果熟最早是多花黄精,最晚是滇黄精,相差9 天;滇黄精最早进入枯萎期,黄精则最晚,相差6 天。蒋燕锋等[28-29]发现17 个不同种源的多花黄精地下茎增产幅度几乎都高于黄精和滇黄精(如表1 所示);多花黄精根茎的多糖含量在9.70%~36.32%之间,平均为22.21%,是《中华人民共和国药典》规定值7%的3.17 倍。较高的产量和有效成分含量是多花黄精在3种基源植物中人工种植最多的原因。
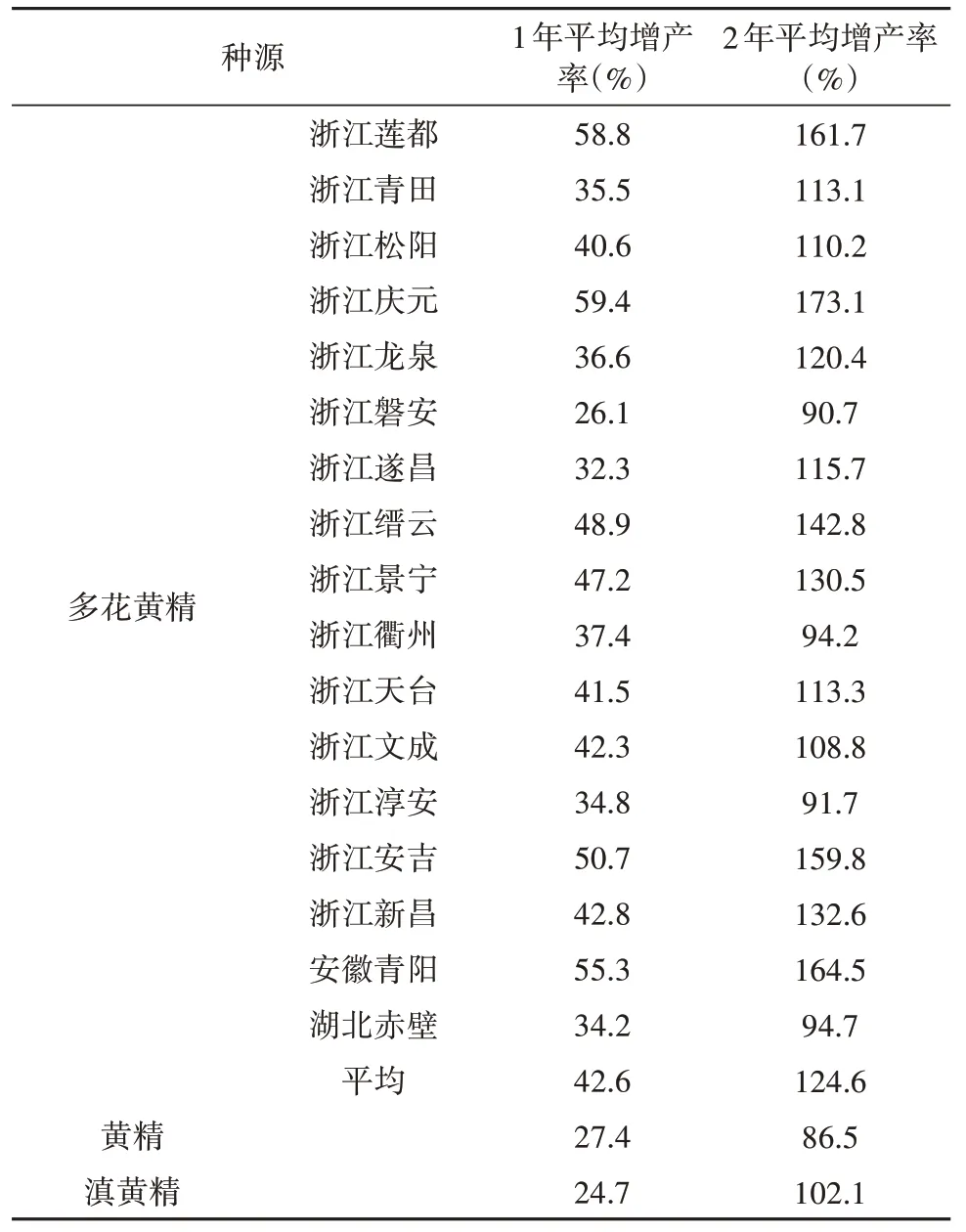
表1 黄精、滇黄精及不同种源多花黄精的生长量Table 1 Increment of P. sibiricum,P. kingianum and 17 P.cyrtonema provenances from different source regions
3 种不同基源物种根状茎在有效成分的种类上同样表现出巨大的差异。红外光谱检测发现滇黄精甾体皂苷类成分最为丰富,黄精、多花黄精中皂苷含量较低;黄精中糖类成分含量最少[30],可根据收获目标成分选择相应的物种来种植。黄精基源植物在糖类成分的种类上也存在较大差异。多花黄精的多糖样品结构均含有阿拉伯糖、半乳糖、葡萄糖、甘露糖和半乳糖醛酸,黄精多糖仅由单一果糖构成,滇黄精多糖则主要由葡萄糖组成[31-34]。通过多元回归方法对UPLC-MS 检测所得的代谢物图谱进行分析,根据测得的所有代谢产物种类及含量,可将供试药材中3 种法定基源黄精明显聚为3类[35]。进一步研究表明,相较于滇黄精和多花黄精,黄精中缺失盐酸氨基葡萄糖;多花黄精中D-甘露糖较滇黄精和黄精量少[36]。
即便同一物种、不同种源的3 种黄精基源植物在农艺性状及有效成分含量方面也具有很大差异。周新华等[37]分析了来自不同地区的9个多花黄精野生种群的12个表型指标,发现不管是在种群间还是在种群内,除果实横纵比外,野生多花黄精叶长、叶宽、叶片长宽比、叶面积、全株叶面积、地径、株高、果实纵径、果实横径、种子千粒重都有显著差异,各表型性状的平均变异系数为12.16%,变异幅度范围为4.76%~23.63%。涂明锋等[38]对8 个产地收集的多花黄精新鲜根状茎成分进行对比,江西宜春的多花黄精多糖含量最高,为14.77%,湖南湘西的仅为2.66%;湖南张家界的多花黄精黄酮含量最高,为1.09 mg/g,广西桂林的仅为0.35 mg/g;四川广安的多花黄精多酚含量最高,为1.45 mg/g,江西宜春的仅为0.47 mg/g。王丹等[39]比较了5 个不同产地的多花黄精根状茎有效成分,多糖含量在8.09%~11.78%范围内变化,以湖南娄底地区的最高,总皂苷含量范围为2.73%~5.01%,以湖南永州地区的最高;总黄酮含量范围为0.21%~0.71%,以湖南张家界地区及四川甘孜地区的较高。上述研究结果为我们选择优良黄精种源提供了参考。
1.3 黄精基源植物种质的遗传多样性
形态特征和农艺性状表明不同种质,不同种源的黄精存在丰富的遗传变异。早期研究多集中在细胞学层面,通过染色体数目和形态分析黄精属植物的遗传多样性。目前黄精属60多种植物中,做过染色体研究的有47 种左右,总共222 条染色体计数,已知其中14种具有不同的染色体数目,3种黄精基源植物中,多花黄精和滇黄精都发现了种内染色体基数变化,已发现的多花黄精的染色体基数X=9、10、11、12,滇黄精X=13、15、16,目前发现的黄精种质染色体则皆为X=12[40]。黄精属植物的核型在不同居群间及同一居群中都存在多样性(见表2),同一物种中,每份资源的核型公式也几乎都不相同[41]。通过核型分析,可对同一物种不同居群的区分起到一定的作用,同时也可助于发现新物种,如金孝锋等[43]将核型分析与形态分析结合,发现了浙产多花黄精的一个染色体基数X=12 的变种古田山黄精。
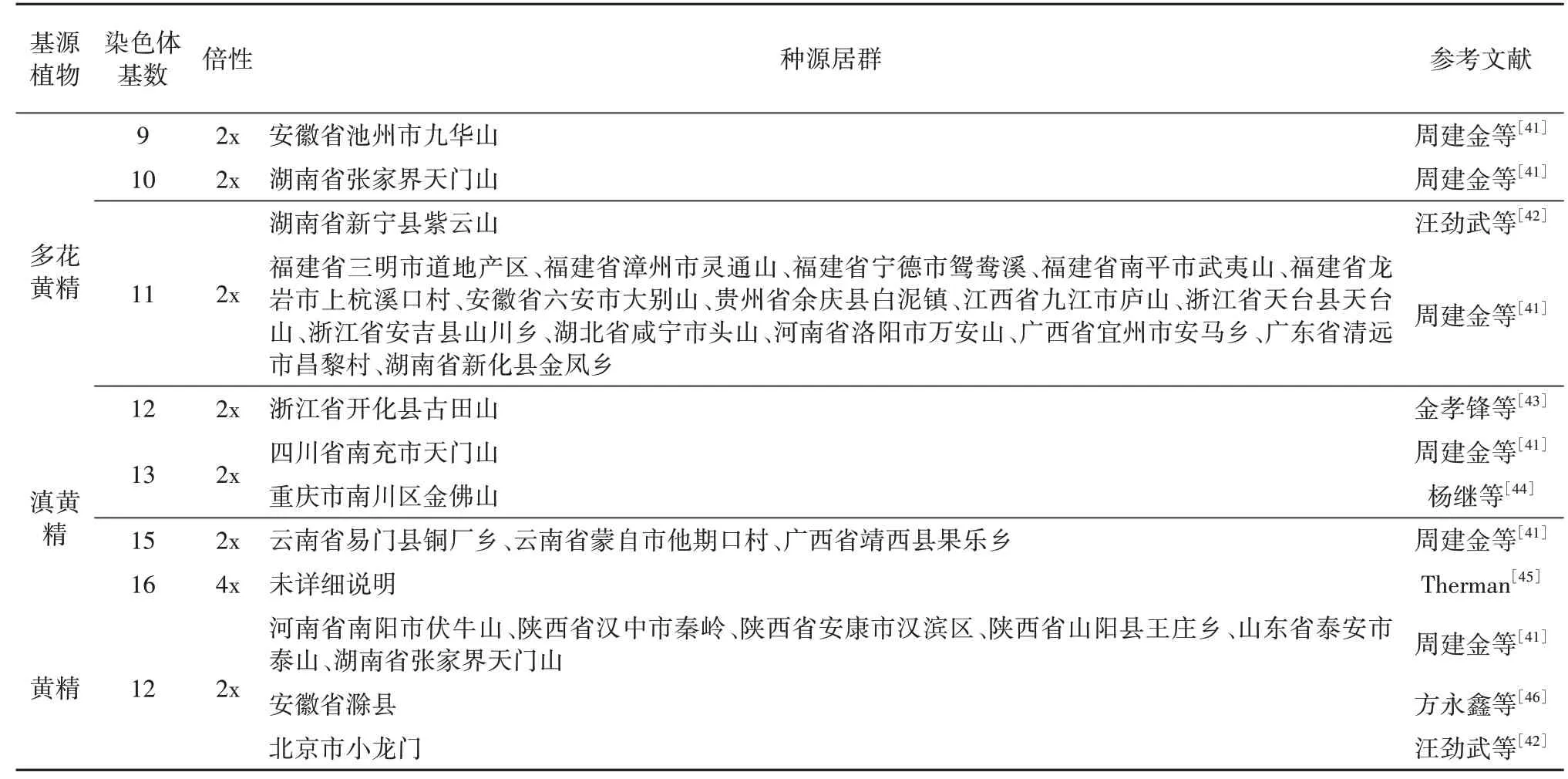
表2 各黄精基源植物的染色体信息Table 2 Chromosome information of origin plants species of Polygonati Rhizoma
黄精属植物在自然条件下主要是通过营养体繁殖,因此更易产生染色体组型异化,且黄精属植物的染色体变异为非整倍性变异,无论是种间还是种内都存在较大的变异,其形态学上界定的种在其遗传上并非同质[47]。目前黄精属内物种的分类和物种鉴定仍存在一定困难,如湘黄精曾经作为独立物种报道,但此后形态学比较和分子系统学表明,湘黄精应与滇黄精属于同一物种[48-49]。
随着分子生物学技术的发展和进步,基于DNA克隆和序列分析的分子鉴别技术在遗传多样性分析方面发挥了重要作用。Tamura 等[50]最早将DNA条形码技术应用于黄精属植物分类研究,此后,分子生物学技术被广泛应用于揭示多花黄精遗传多样性及地理分布特征。李巧玲等[51]采用ISSR 分子标记技术对黄精属8 个种的植物进行多态性分析,其多态性位点百分率达到96.8%,植物间遗传相似性较低,具有丰富的遗传多样性。张恒庆等[52]采用ISSR分子标记技术分析了大连地区3个黄精种群,5个多花黄精,黄精种群多态位点比率86.86%,遗传分化系数为0.398 2,多花黄精种群多态位点比率94.41%,遗传分化系数为0.308 4,黄精和多花黄精天然种群均具有较高的遗传多样性,且多花黄精多样性高于黄精。张红梅[53]采用ISSR 分子标记技术分析了安徽省内分属于4 个物种的14 个黄精属植物群体,根据产生的条带聚类,同一种源的种质聚为一类,其中5 个黄精居群遗传距离在0.414 2~0.496 4,4 个多花黄精居群遗传距离在0.388 2~0.555 3,所检测的4 个黄精物种整体遗传多样性为0.350 0,种间分化系数为0.432 8。小范围的分子标记聚类基本上符合传统分类学方法,但在更大范围的研究中则出现了种间交叉及地理分布交叉现象,如朱巧等[54]对14个省38个县市收集的60份材料黄精属种质资源进行了SSR 遗传差异分析,发现在遗传距离阈值为0.26 时可聚为4 大类群,大部分同一物种或近缘物种的材料聚为一类,如10份滇黄精材料聚为一类,但有6 份材料并没有和同种的其他材料聚在一起,如7份黄精材料与卷叶黄精聚为一类,但也有3份黄精材料与27份多花黄精材料和3份玉竹材料聚为一类,另有3 份多花黄精材料则与湖北黄精聚为一类,群体遗传结构分析则表明,其中50份材料遗传组分相对单一,但有10份材料渗入了其他群体的遗传信息,表明不同类群间存在基因交流,黄精属内物种的界限相对模糊。将60份种质按照地理来源划分,其遗传多样性指数从大到小依次为西部地区>华中地区>华南地区>华东地区,西部地区的遗传多样性较其他地区高,可能是我国黄精属植物的起源中心。
大范围的遗传多样性研究同样揭示了黄精物种内地理分布与遗传分化的关系。籍蓉蓉[55]采用SSR 分子标记技术对安徽省13 个多花黄精居群进行分析,发现所收集的种质具有中等以上的遗传多样性,略高于多年生植物平均值,结合部分形态数据,可将13 个多花黄精居群分为3 类,宣城、池州市的6 个居群为一组,黄山市的4 个居群为一组,大别山区的3 个居群为一组,与山脉地理分布一致。刘新等[56]采用ISSR 分子标记技术对浙江、安徽、江西、福建、湖南、湖北6个省20个种源的118份多花黄精种质进行综合分析,发现平均多态性百分率为94.62%,种源内遗传多样性在33.85%~60.00%,UPGMA 聚类分析显示,同一种源的种质基本上聚为一类,说明种源间遗传分化大于种源内遗传分化;在遗传相似系数等于0.61 时可将种质聚为武夷山脉、武夷山脉和罗霄山脉、大别山脉、洞宫山脉和天目山脉4类,与地理区位分布一致,表明山脉之间的平原、水域隔离是群体间遗传分化的主要原因之一。
2 黄精基源植物种质多样性的开发利用
2.1 黄精基源植物基因功能研究方面的利用
植物种内所有个体的所有遗传性状的基因构成了该物种的基因库,其中可能蕴藏着丰富的已知或未知的有用基因。种质资源遗传多样性的研究将为评估基因资源的开发前景提供了重要的信息,而关键基因的发现和利用将是育种取得突破性成就的关键。关于黄精基因方面的研究起步较晚,目前尚无可参考的完整基因组测序结果,仅有叶绿体基因组序列报道。Pan 等[57]2020 年已完成黄精叶绿体基因组的测序,滇黄精的叶绿体基因组由Jin等[58]于2020年完成测序。黄精属内近缘物种卷叶黄精、玉竹以及一个尚未确定分类地位的冬季不落叶黄精属物种(与黄精的遗传距离远于轮叶黄精,但近于狭叶黄精)也已有完整的叶绿体基因组序列可供参考[59,61],有助于黄精属内物种的分类关系研究和种源鉴定。
目前对3种基源植物基因功能的研究多采用解析过程不需要参考基因组的RNA-Seq 方法,且主要集中在研究黄精基源植物的次生代谢产物合成途径方面。黄酮、多糖和甾体皂苷是黄精药材中药理作用较为明确的主要药用成分,3 种黄精基源植物都已有相关的转录组研究结果报道。而基于遗传转化体系的黄精基因功能验证则由于3种黄精基源植物都未建立完整的遗传转化体系而尚未开展,黄精的有效成分积累及其与环境因素的互作机制尚不完全清楚。
2.1.1 黄精基因功能的研究
党凯凯等[62]以皂苷、多糖含量差异都较大的河南洛阳产黄精和陕西略阳黄精为材料进行高通量转录组测序,经注释分析其中146条Unigene 对应到甾体皂苷生物合成途径中的32个相关酶,几乎覆盖了该途径上的所有节点,其中10 个酶对应的10 条Unigene 表达在两个品种间具有显著差异,其中8 个酶对应的合成基因表达量变化规律在不同产地黄精和黄精不同部位中与甾体皂苷积累情况相似,表明这些基因可能在黄精的甾体皂苷合成中起关键作用;王世强[63]发现138条Unigene 对应到黄精多糖生物合成相关的酶,其中8个酶对应的88条Unigene在两个品种间具有显著差异,其中5 个酶对应的合成基因表达量与多糖积累在不同种质间正相关,己糖激酶合成基因与多糖积累负相关,β-呋喃果糖苷酶、己糖激酶和甘露糖-1-磷酸鸟苷酰基转移酶是多糖积累的关键酶。
2.1.2 滇黄精基因功能的研究
肖韵铮等[64]以根、茎和叶为样本进行转录组测序,发现器官间差异表达基因主要集中在核糖体、光合作用、碳代谢、苯丙烷类生物合成、淀粉和蔗糖代谢等过程,表明与类黄酮合成相关通路是差异基因主要富集通路之一。结合相应器官的类黄酮含量,筛选出与类黄酮生物合成相关的10个差异表达基因,包括查尔酮合成酶(CHS)、查尔酮异构酶(CHI)、二氢黄酮醇-4-还原酶(DFR)、类黄酮3′-羟化酶(F3’H)、香豆酸-3′-羟化酶(C3’H)、咖啡酰辅酶A 甲基转移酶(CCoAOMT)、无色花色素还原酶(LAR)、反式肉桂酸酯-4-单加氧酶(CYP73A)、花青素还原酶(ANR)和黄酮醇合成酶(FLS)。其中,查尔酮合成酶和查尔酮异构酶基因的表达量与类黄酮含量呈显著正相关,可能决定了类黄酮的合成。
2.1.3 多花黄精基因功能的研究
祝明珠[65]对多花黄精根、根状茎、茎及叶片的转录组测序,发现263个Unigene 对应到多糖合成途径中的14 个关键酶,薯蓣皂苷合成途径中则有4 种关键酶检测到对应的Unigene 共39条。单春苗等[66]以类似方法检测到皂苷合成途径中12 个关键酶对应的共97 个Unigene,涉及皂苷合成的Unigene 达到502 条,且其中35 条在根状茎中特异性表达。廖荣俊[67]对多花黄精幼苗期根状茎进行转录组测序中发现27个皂苷合成相关酶对应的共计113个Unigene,其中45 个与拟南芥中的酶同源,为多花黄精的功能基因组学研究提供了丰富的参考数据。祝明珠等[65]、单春苗等[66]进一步克隆了甾醇合成途径中第一个关键酶环阿屯醇合成酶的完整序列,并通过序列分析预测了其蛋白结构,为进一步研究其表达调控奠定基础。CIGR(Chitin-inducible gibberellinresponsive genes)和SCL21(SCARECROW-LIKE 21)属于响应赤霉素信号的GRAS 转录因子家族中的PAT1亚家族,该家族基因在光敏色素的转导方面具有重要作用。为探究多花黄精无菌快繁过程中环境条件的影响,吕煜梦[68-69]克隆了多花黄精中的Pc-CIGR 和PcSCL21,发现两个基因的表达产物都定位于细胞核,具有器官表达特异性,均在叶中表达量最高,PcCIGR 基因表达受蓝光和短日照诱导,Pc-SCL21基因表达受蓝光、绿光和长日照诱导,两个基因在激素处理下表达量和薯蓣皂苷含量呈现显著正相关关系。长梗黄精是多花黄精的易混品,在形态及地理分布上与多花黄精极为相似,在多花黄精供不应求的环境下具有作为候选替代种的潜力,徐惠龙等[70]以种源收集自同一地区,栽培条件一致的多花黄精和长梗黄精幼嫩根状茎为样本进行转录组测序,发现多花黄精与长梗黄精在多糖和薯蓣皂苷相关代谢通路表达差异较少,228 个Unigenes 编码3 条多糖生物合成途径中13 个关键酶,但只有35个Unigenes 有显著性差异,64 个Unigenes 编码薯蓣皂苷生物合成途径9个关键酶,仅6个Unigenes有显著性差异,为扩大黄精药材的植物资源提供了参考依据。
2.2 在黄精基源植物遗传育种方面的利用
药用植物育种的主要目的是获得有效成分含量高的品种。任何一个新品种的培育都是在大量的植物资源基础上通过选择、杂交、回交、诱变等方法修饰、加工、改良后培育出来的。由此可见种质资源及其遗传性状分析是育种的基础和关键。
黄精属内形态学上界定的种在其遗传上并非同质,同样,遗传上近缘的黄精属植物同样可能存在较大的表型差异。周新华等[37]对生长在野外的多花黄精种质进行表型聚类分析,发现多花黄精种群内的变异(36.24%)大于种群间的变异(15.37%),多花黄精种群内的遗传分化较种群间的遗传分化更容易实现。王旭军等[71]收集不同种质进行人工栽培,发现大部分农艺性状与药用有效成分含量的遗传方差均小于其环境方差,遗传变异系数也均小于其环境变异系数,除株高、根茎多糖含量外,其他性状的遗传力均比较低。吴媛媛[47]将不同种质于同一环境下栽培,发现不同多花黄精种质的株高、叶宽、果实直径和百粒重等性状的遗传力较高,分别为71.03%、57.156%、63.32%和77.83%,且其变异系数均低于10%,表明这些性状不易受栽培环境条件影响,但根长、根平均直径、根最大直径、根最小直径等根部形态性状的遗传力普遍偏低,均低于30%,且其变异系数较高,表明根部形态性状受栽培环境条件的影响较大。黄精基源植物遗传性状分析对其遗传育种具有很好的指导意义。
黄精基源植物主要依赖营养繁殖且相关研究起步较晚,目前品种选育方面成果较少,但已有多花黄精品种“皖黄精3 号”和“丽精1 号”面世。“皖黄精3 号”由安徽省农业科学院选育而成的。彭星星等[72]从安徽旌德周边山中收集大量的野生多花黄精资源,以系统选育的方式从中筛选出9 个各有特色且表现一致的类群,在此基础上,对根状茎形态及产量、相关成分含量、病害发生情况等进行考察和分析,进一步筛选出品种“皖黄精3 号”。该品种具有对根腐病的高度抗性,且株高可达到95 cm,年平均生长期125 天左右,栽培3 年单株可产根状茎395.7 g,黄精多糖含量13.06%,为健壮高产的优良品种,于2019 年11 月通过安徽省非主要农作物品种鉴定登记。“丽精1 号”为丽水市林业科学研究院选育,刘跃均等[73]从采集自安徽省青阳县酉华镇多花黄精群体中,以系统选育的方式经优选扩繁而成。该品种适宜种植于浙江省海拔800 m 以下地区,其株高84.2~125.5 cm,地上部分年生长期200天左右,根茎多糖含量14.61%~14.73%,明显高于栽培的滇黄精(多糖含量6.70%~14.28%)、野生多花黄精(多糖含量7.21%~11.47%)以及不同产地野生黄精(多糖含量6.34%~16.79%)。
目前黄精基源植物的育种仍以在野生种质收集的基础上系统选育为主,且由于其遗传多样性,在一段时间内野生资源的收集与系统选育仍将是黄精基源植物遗传育种的主要方式。为了更好地评价、筛选黄精种质资源,焦劼等[74]初步编制了《药用植物种质资源鉴定评价技术规范黄精》,筛选了涉及农艺学、生态学、生药学方面的60 个性状用于黄精种质资源的鉴定,建立了包含其中21个性状的多目标决策分析模型用于黄精种质资源的评价,并对评价方法和技术进行了规范。依据该技术规范检测了43 个黄精基源植物种质,其中,产自陕西略阳、陕西安康、甘肃陇南的黄精,产自重庆武隆、安徽泾县、广西贺州、广西宜州及重庆綦江的多花黄精种质既是优良的遗传种质材料,在陕西略阳地区的种植环境下也是优良的药用植物品种。在此基础上,焦劼等[74]进一步尝试通过逐步距离压缩法筛选黄精基源植物核心种质,从地理分布不同的9 个居群,共47 份种质材料中筛选出13 份作为核心种质,保留的遗传多样性在多种评价标准下均达到90%以上,为引种和遗传育种初步建立了基础。
3 总结与展望
随着我国居民生活水平的不断提高,对于健康的重视程度不断提升,中医药学在慢性病、亚健康的医疗调理及养生保健方面的作用,特别是在新冠病毒防治方面的作用获得了人们的广泛认可。中药材是中医药事业和大健康养生产业发展的重要物质基础。中药材传统上主要来源于野生资源采挖,产量已不能满足日益增长的需求,缺乏保护意识导致药材野生资源枯竭,虽然经过多年的发展,我国中药材产业得到了快速的发展,但仅有部分药材种类具有相对成熟的生产和评价体系。对于黄精这类基源植物分布广,遗传变异和环境变异大,生长周期长,因依赖营养繁殖导致繁殖系数低、品种退化的药材,其规范化栽培、种质资源保护与开发利用、药材质量控制方面仍然存在较大问题,对于其有效成分积累与环境互作的机理机制仍不清楚。
长期以来对黄精的研究集中于有效成分的药理机制,对黄精本身的形态生理及遗传方面的研究较少,制约了黄精种植产业的发展。近年来,随着分子生物学技术及代谢产物检测技术在黄精研究上的应用,黄精的种质资源评价和开发方面有了一定的发展,但由于相关研究起步较晚,仍存在很多不足之处。在基础研究方面,主要是基因组测序以及遗传转化体系的空白。目前3种黄精基源植物尚没有完整基因组数据可供参考,导致转录组测序获得的数据分析受到限制,Unigene 数据率依然较低,亟需在基因组测序方面取得进展,且3 种黄精基源植物都未建立稳定的遗传转化体系,导致转录组获得的具有增产潜力的基因难以在黄精自身上进行功能验证,这严重限制了黄精基因功能的研究和利用。在遗传资源应用方面,仍要加快黄精基源植物的种质筛选及品种选育。此前黄精种质收集及筛选工作分散且进展缓慢,仅仅只有2 个多花黄精品种经过品种认定,但目前黄精基源植物的种质评价技术规范以及核心种质筛选技术已经初步建立,这一标准的应用有望大幅提高黄精种质资源筛选及品种选育的效率。此外,王世强等结合转录组、代谢组和分子标记技术以及现有QR 软件建立了黄精DNA 指纹图谱、种源二维码和相应的数据库,极大地方便了种质信息的收录和查询[75-76],此类多种生物技术联合、生物技术与软件开发结合将是未来的主要发展方向。
