环境未培养微生物中新型抗生素的发掘研究进展
2022-04-15王梦歌彭金金戢博阳纪晓俊魏勇军
杜 瑞,王梦歌,彭金金,戢博阳,纪晓俊,魏勇军
(1.郑州大学 药学院 郑州大学合成生物学实验室 药物关键制备技术教育部重点实验室,河南 郑州 450000;2.查尔姆斯理工大学 生物与生物工程学院,瑞典 哥德堡 SE-41296;3.南京工业大学 生物与制药工程学院,江苏 南京 211800)
抗生素是临床治疗病原微生物感染的最常用药物,但由于已有抗生素的广泛应用,甚至滥用导致微生物产生耐药性,使现有抗生素的疗效降低甚至无效;耐药性致病菌引起的感染已成为全世界面临的最主要健康问题之一[1-2]。抗生素是微生物的次级代谢产物,筛选并表征微生物中的抗生素合成基因簇是解决致病菌抗生素耐药性的有效途径之一。自20世纪70年代以来,传统培养分离微生物并筛选抗生素的方法已很久未发现新化学结构的抗生素[3]。近年来,环境未培养微生物成为抗生素发掘和研究的重点关注对象,因此,本文综述近年利用新技术发掘环境未培养微生物中新型抗生素的方法,以期为新型抗生素的发现及规模化生产提供参考。
1 新型抗生素有较大需求
1.1 耐药致病菌严重危害人体健康
美国疾病控制中心的报告显示美国每年约有280万人发生耐药致病菌感染,约3.5万人死于耐药细菌感染(https:∥www.cdc.gov/drugresistance/index.html)。若无新的治疗方案,普通的细菌感染即可对人类构成致命的威胁。据估计,全球因抗生素耐药致病菌引起的人类感染在2050年将增加10倍,将导致超过1 000万人的死亡,抗生素耐药菌对人类产生的危害将会超过癌症[4]。
我国是抗生素的生产和使用大国,每年人均消耗抗生素138 g左右。耐药性细菌在我国分布广泛[5-6]。中国细菌耐药监测研究报告指出,2017—2018年,我国临床分离的表皮葡萄球菌(Staphylococcusepidermidis)中,对甲氧西林有耐药性的菌比例高达85.7%,大肠杆菌(Escherichiacoli)中超广谱β-内酰胺酶的检出率也达54.6%[7]。此外,其他多种病原菌的抗生素耐药性检出率也较高[8]。普通抗生素对耐药性细菌的疗效不明显,患者感染后难以治疗痊愈,曾被有效治疗的细菌感染性疾病可能再次对人类构成致命威胁[9]。当前,除减少抗生素滥用并控制抗生素耐药基因的进化、传播与扩散外,还迫切需要开发新型抗生素对抗耐药细菌[10]。
1.2 可培养微生物已难以发现新型抗生素
抗生素主要分为聚酮类化合物和非核糖体多肽类化合物[11],其中,聚酮类化合物(polyketides)通常由酰基辅酶A活化的底物之间的重复脱羧缩合合成[12];而非核糖体多肽类化合物(non-ribosomal peptide,NRP)是由非核糖体多肽酶催化相应的单体氨基酸合成。聚酮化合物的骨架合成元件包括聚酮合成酶(polyketide synthase, PKS)、酰基转移酶、酰基载体蛋白及脱水酶、烯醇还原酶、糖基转移酶等修饰元件[13];非核糖体肽类化合物的骨架合成元件包括非核糖体肽合酶(non-ribosomal peptide synthase, NRPS)、肽酰基载体蛋白、氨基酸腺苷化结构域、缩合结构域以及硫酯酶等修饰元件[14-15]。这些抗生素合成基因通常成簇存在,长度可达上百kb,由多个相似性较高的抗生素骨架合成基因及糖基化、甲基化等修饰基因组成[12,14]。
现有抗生素主要来源于可分离培养的细菌,其中一半以上的临床应用抗生素来源于放线菌门(Actinobacteria)链霉菌属(Streptomyces)的微生物[2, 13]。目前放线菌门中的抗生素资源已被深度挖掘,所以利用可培养微生物发酵筛选到的抗生素通常与已知活性的抗生素相同或相似,因此科研人员已难以从可培养微生物中筛选到新型抗生素[11]。自20世纪80年代以来,已近40年没有新型结构或作用机制的新型抗生素上市。目前,耐药细菌对被化学结构修饰得到的抗菌药物敏感度不强,所以难以利用结构修饰从已知结构的抗生素中筛选到满足临床使用的新型抗生素[2]。由于抗生素特别是源于放线菌中针对革兰氏阴性菌类的抗生素被“过度”挖掘,同时难以从化合物库中筛选到新型抗菌物质,所以亟须开拓其他新的抗生素发掘策略。
2 微生物组学在新型抗生素发掘中的应用
分子微生物学研究表明,环境中可能存在着数百万种微生物,但培养分离技术手段有限,大部分的微生物是未培养的[16],这是难以从可培养微生物中筛选到新型抗生素的根本原因。各种环境中蕴藏的独特的、未挖掘的、多样性丰富的微生物及其天然产物,是新型抗生素的潜在来源[17]。抗生素作为微生物次级代谢产物,其合成受到环境等因素的严格调控,在环境中抗生素合成基因通常不表达或低表达,导致很难直接从环境微生物特别是未培养微生物中筛选到新型抗生素[18]。因此,挖掘并表征环境未培养微生物中抗生素合成基因簇是发掘新型抗生素的重要途径。
随着高通量测序技术[19]的发展,不依赖于培养的微生物组学技术,尤其是宏基因组学技术[20]逐渐完善并被广泛应用。宏基因组学是以环境样品中的微生物群体基因组为研究对象,利用测序分析和功能基因筛选为研究手段,阐明微生物多样性、种群结构、进化关系、功能活性、相互协作关系及与环境之间的关系为研究目的的微生物学研究方法[21]。通过宏基因组学研究,能够有效精准地鉴定出不同微生物群落结构和功能的差异,利用多个抗生素合成基因/元件/模块组合分析预测抗生素结构和功能的多样性和新颖性,有目的筛选抗生素合成基因簇。此外,还可以通过对Cosmid等基因文库的筛选,获得抗生素合成基因簇,进而获得新型抗生素[22](图1)。如,通过对环境中未培养微生物进行高通量筛选,发现了破坏细菌细胞壁的抗生素——Teixobactin[23]。
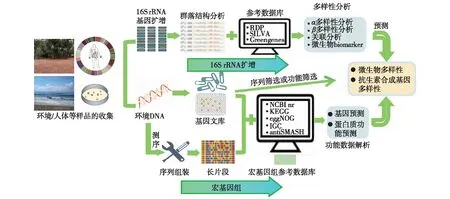
图1 宏基因组学等技术及其在抗生素合成基因簇发现中的应用[22]
2.1 微生物多样性与抗生素合成基因多样性的解析
各种环境中的未培养微生物是挖掘新型抗生素的重要来源。目前,已有多位学者对源自土壤、海洋及人体等的微生物组中的抗生素合成基因簇进行了解析[24-26]。
土壤是多数抗生素的主要环境来源[27]。现阶段分离可培养微生物得到的抗生素均为已知的[28],利用适用于未知物种、未知基因检测的非培养依赖的微生物组学方法揭示环境微生物及其天然产物合成基因,挖掘出大量未知的新型天然产物合成基因簇,为新型抗生素的发现提供基础[17, 29]。Brady团队的Hover等[30]采用宏基因组测序结合细菌人工染色(BAC)文库筛选策略,从近2 000个土壤样本中发现了1种未知抗生素合成基因簇,并利用该基因簇合成了耐甲氧西林金黄色葡萄球菌(Staphylococcusaureus)等多药耐药致病菌的新型抗生素Malacidins。Charlop-Powers等[31]研究发现,在美国纽约等都市区的土壤中存在着多种新颖的抗生素合成基因簇,且与其他自然环境中的天然产物合成基因/基因簇明显不同。Crits-Christoph等[17]对美国加州的草地土壤大规模宏基因组学测序发现了1 159个新颖的天然产物合成基因簇;这些基因簇不仅来自已知的传统产抗菌如放线菌门和变形菌门(Proteobacteria),还有一半以上来源于酸杆菌门(Acidobacteria)(来源最多)、疣微菌门(Verrucomicrobia)、Rokubacteria和Gemmatimonadetes,其中,对酸杆菌门微生物测序后发现的天然产物合成基因簇是之前从未发现过的,这为发现新抗生素提供了基础。
海洋微生物中也分布着丰富的抗生素合成基因簇[26]。深海的环境恶劣、营养物质供给匮乏,深海环境中的微生物有可能通过产生特定的次级代谢产物抑制周边微生物的生长,以争夺对自身有利的生存空间[32]。受到这一化学生态学原理的启发,同时海洋环境微生物的研究相对较少,近期科研人员将发掘新型抗生素的目光转向海洋。Jackson等[33]研究发现,海洋微生物中含有丰富的编码聚酮类化合物、非核糖体多肽、细菌素(Bacteriocins)、兰蒂肽(Lassopeptides)和拉索普肽(Lantipeptides)等代谢产物的合成基因簇,其中深海微生物编码非核糖体多肽的基因簇多样性较高。Zhang等[34]利用微生物组学方法筛选海鞘(Pyrosomellaverticilliata)的微生物组后得到具有强抗菌活性的天然产物Turbinmicin。海绵(Porifera)是一种底栖滤食动物,大量的微生物寄存于其独特的滤食系统中,在浅水和深海海绵体内中分离出的微生物次级代谢产物对多种临床相关病菌和酵母菌均表现出独特的抗菌活性[35]。Jackson等[33]通过宏基因组学测序研究4个不同进化方向的深海海绵衍生链霉菌属的次级代谢产物合成基因簇,结果发现,海绵中含有丰富多样且高度新颖的次级代谢产物合成基因簇。Wei等[36]对黄海海底沉积物的研究发现,海洋中的大量抗生素合成基因簇与已知的抗生素合成基因簇同源性在50%以下;通过序列筛选,从构建的Fosmid克隆文库中获得10个新的抗生素合成基因簇。此外,Uzair等[37]从海藻微生物群落中分离到对耐药菌有抑制作用的潜在的新型抗生素。鞠建华研究团队的Zhu等[38]从深海微生物群落中提取出对革兰氏阳性菌和厌氧性革兰氏阴性菌具有高效抑制效果的含有氨基己糖结构的核苷类抗生素A201A。Ma等[39]从深海放线菌中提取分离出具有明显抑制结核分枝杆菌活性的结构新颖的抗生素怡莱霉素(Ilamycin C)。这些研究还发现,即使是相邻海域的微生物,其种类和抗生素合成基因簇也显著不同,这增加了从海洋环境中发现新型抗生素的可能性。
人体内具有丰富多样的微生物群落[40]。每个人体内的微生物种类高达上千种,而且不同个体之间、同一人的不同部位之间的微生物群落均有差异[41]。人体胃肠道微生物菌群一直是抗生素研究的关注重点,宏基因组研究表明在全球各地人类肠道中存在着丰富多样的未培养菌种[42]。人体内同样蕴藏着丰富的抗生素合成基因簇,是发现新型抗生素的重要来源。Pasolli等[43]利用宏基因组学方法从一些未知的微生物种群中识别出数千个微生物基因组,丰富了人体相关微生物的基因组,为从人体微生物群落中发现新型抗生素拓展了来源。Zipperer等[44]在人体鼻腔微生物中发现了能够抑制耐药金黄色葡萄球菌的抗生素Lugdunin。Sugimoto等[45]利用MetaBGC策略对人体微生物组中的抗生素合成基因簇进行发掘后发现,不同地区和不同菌株的微生物具有不同的抗生素合成基因簇,且其中包含多个聚酮化合物。Imai等[18]对动物线虫体内肠道菌Photorhabdus中天然产物合成基因簇的解析及异源表达,获得了新型抗生素达罗布汀(Darobactin);经测试发现,达罗布汀对抗耐药性革兰氏阴性菌具有较强的抑菌活力。除序列筛选外,利用功能基因筛选的方法也能从环境微生物组中获得大量新型的抗生素合成基因簇,但是该方法较难发现完整的基因簇且工作量大,目前已较少使用[41]。
由此可见,在土壤、海洋、人和动物体等环境中蕴含着丰富的新颖抗生素合成基因簇,是新型抗生素的重要环境来源。但多数抗生素合成基因簇表达水平较低或是不表达,需开发新的抗生素合成基因簇表征方法。微生物组学的发展为发现新型抗生素合成基因簇提供了基础,通过开发新方法,鉴定表征以“唤醒”这些“沉默状态”的抗生素合成基因簇,有望发掘出应用于临床的新型抗生素[46-47]。
2.2 生物信息学及多学科交叉为新型抗生素的发现奠定基础
微生物组学的发展已产生了海量的数据,为抗生素合成基因簇的鉴定表征奠定了基础。为了从海量的数据中快速发掘新型抗生素合成基因簇,目前科学家已开发出多个抗生素(天然产物)合成基因簇数据库,建立并改进了抗生素合成基因簇的预测方法。Helfrich等[48]开发出预测反式乙酰转移酶聚酮化合物的重要工具TansATor,并利用该技术挖掘到数个新颖的抗生素合成基因簇。Blin等[49]开发了预测抗生素和次级代谢产物合成基因簇可能合成产物的工具antiSMASH。Navarro-Muoz等[50]基于生物信息学手段构建了一种新计算工具BiG-SCAPE,该工具可以对微生物群落进行分析并快速挖掘其中的生物合成基因簇。这些研究为科研人员开发可用抗菌化合物打下了坚实的基础,同时生物信息学在基因片段的功能开发、蛋白质结构预测、药物结构设计、新药研发及合成中起着至关重要的作用,可加速新型抗生素研发进程[51-52]。
目前,已经发现的用于合成非核糖体多肽的稀有氨基酸种类多达300个,种类繁多的稀有氨基酸使得这类代谢产物丰富多样。同时,固相肽技术的迅猛发展加快了多肽合成技术的进步,降低了多肽合成的成本。基于此,Chu等[53]开发出了合成-生物信息天然产物(syn-BNPs)的方法,该方法首先利用生物信息学方法预测天然产物合成基因簇合成的多肽类化合物,然后运用化学方法合成预测的多肽,并在体外验证其活性,以此获得了新型抗生素Humimycins。Fritz 等[54]利用研发的新型生物信息学工具NRPPUR成功从人肠道菌的基因组筛选到89组新型的抗生素合成基因簇。加拿大科学家Culp等[55]通过解析土壤细菌的生物合成基因和抗性决定簇,预测出1个糖肽家族生物合成基因簇,以此获得一种具有新功能机制的潜力抗菌化合物——Corbomycin。由此可见,生物信息学结合化学合成是新型抗生素发现的重要途径[51]。从环境中筛选出合适的抗生素合成基因簇,然后预测它们可能合成的抗菌化合物;选择新颖的、有临床使用价值的抗生素进行化学合成,并进行抑菌实验等功能验证;在此基础上,将有功能的抗生素合成基因簇在模式微生物中进行异源表达,有望大量合成并获得新型抗生素(图2)。
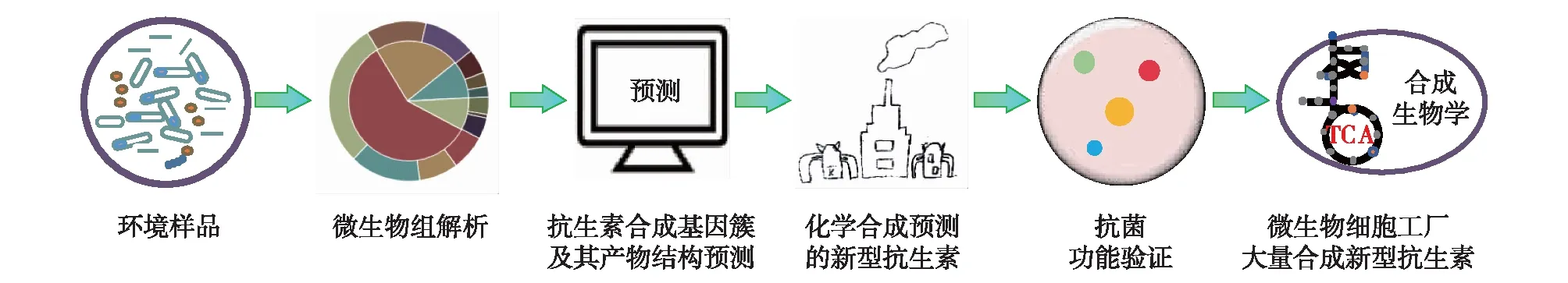
图2 基于基因挖掘生物信息学预测的新型抗生素挖掘与合成方法
人工智能和深度学习技术也被尝试应用于新型抗生素的挖掘[56-57]。Collins团队的Stokes等[56]利用人工智能和深度学习训练的神经网络算法筛选到广谱新型的超级抗生素Halicin。Modongo等[58]通过人工智能和丁胺卡那霉素(Amikacin)暴露来计算耐药菌的耐受浓度,以此推断多重耐药结核杆菌病患者的药物治疗效果,以提高患者的存活率。但是,目前的人工智能等方法还无法大规模筛选环境未培养微生物并对其抗生素合成基因簇进行表征,也无法准确预测抗生素的侧链修饰效应。此外,在化学合成时,对抗生素的侧链进行糖基化、甲基化等功能化修饰很难精确控制。因此,针对复杂未知的抗生素合成基因簇,仍须进行详细表征,并对合成产物的体内/体外抗菌活性和安全性等进行评价。
3 合成生物学在新型抗生素发掘中的应用
合成生物学是近年新兴起的研究领域,是一门综合性交叉学科,主要是以工程学思想为骨架,融合标准化表征的生物学模块,以理性设计为指引,进行模块整合或者从头合成有潜力的、具有目标功能的人造生命的新系统知识和独特理论框架以及特定的功能技术与工程平台[59]。合成生物学已被运用于多个生物制造领域[60],在抗生素的生物合成方面也有着明显的优势[61]。抗生素分子结构多样而繁杂,虽然采用化学方法能够合成少量化合物[53],但是大规模合成仍需在微生物中表达生产(图2)。选择合适的宿主,精准调控抗生素合成基因簇,激活“沉默”抗生素合成基因簇等方法能够有效表征大量新颖的抗生素合成基因簇,用于获得大量新型抗生素[47]。
选择合适的宿主是成功表征抗生素合成基因簇的关键。链霉菌是抗生素的主要来源,也是一种最常用的抗生素合成基因簇表达宿主[46]。目前,临床使用的大多数抗菌化合物由链霉菌合成,如链霉素(Streptomycin)、四环素(Tetracycline)、卡那霉素(Kanamycin)和万古霉素(Vancomycin)等[27]。除已筛选出的抗生素外,链霉菌基因组中还分布着种类丰富的“沉默”天然产物合成基因簇。Fu等[62]利用开发的基于全长RecE的线性-线性同源重组(LLHR)系统,实现了直接克隆DNA 并在异源宿主中表达,获得抗生素Luminmycin。表1总结了部分合成生物技术在抗生素合成基因簇表征中的应用实例。
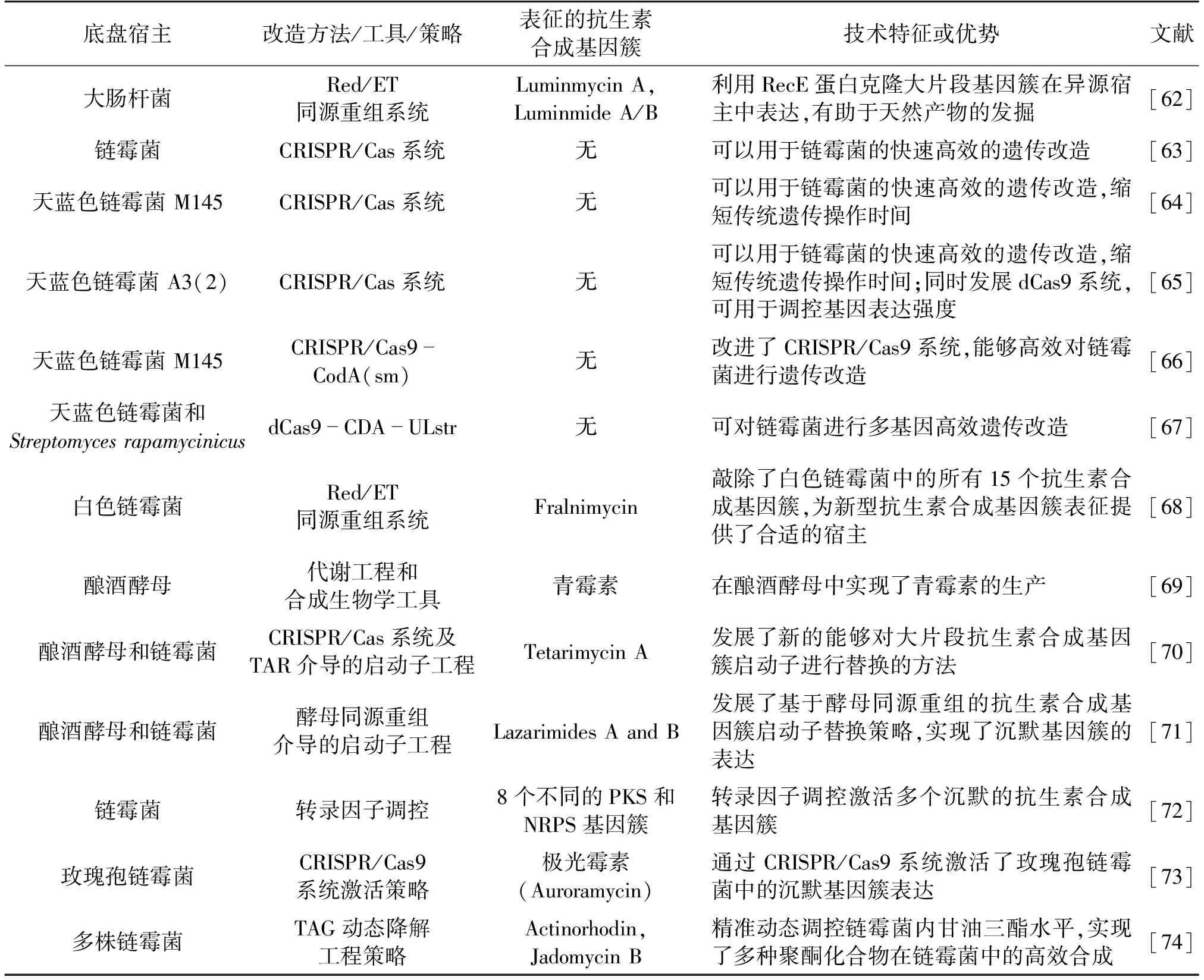
表1 合成生物学技术在抗生素合成基因簇表征中的部分应用
CRISPR/Cas9技术的开发及应用克服了部分天然产物合成基因簇在链霉菌中不表达或表达水平较低的问题,利用CRISPR/Cas9系统对链霉菌等模式宿主工程改造后,显著提升了其表达抗生素合成基因簇的效率[63-65,75]。目前研究人员已研发出多个高效、迅速和可调控的基于CRISPR/Cas9的工具包,用于编辑天蓝色链霉菌,表征抗生素合成基因簇[76]。如,武汉大学孙宇辉团队的Tao等[75]、分子植物卓越中心的姜卫红团队的Li等[77-78]和上海师范大学的芦银华团队的Zhao等[79]开发了基于CRISPR-Cpf1的基因组辅助改造工具,该工具能够精准敲除天蓝色链霉菌中单基因或双基因,效率可达75%~95%,还可提高链霉菌中基因模块编辑和转录抑制的效率,有效加快了链霉菌以及其他放线菌中新颖天然产物合成基因簇的表征以及菌株改造。娄春波团队的Jiang等[80]开发出利用CRISPR/Cas9系统一步克隆大基因簇技术(CATCH),并成功将其用在大片段抗生素合成基因簇如金霉素(Chlorotetracycline)、杰多霉素(Jadomycin)等在链霉菌中的克隆和表达合成。麦克马斯特大学的Culp等[81]以CRISPR/Cas9系统为基础,以启动子工程和转录调节控制为手段,搭建了一个改造链霉菌生产常用抗生素基本组件的底盘细胞平台,能够表征大量的抗生素合成基因簇。Zhao等[67]开发的编辑器dCas9-CDA-ULstr可对链霉菌进行有效的多重基因组编辑,促进了链霉菌菌株的功能基因组和代谢工程研究。Myronovskyi等[68]利用基于BAC库的Red/ET同源重组系统,敲除白色链霉菌中的所有15个抗生素合成基因簇,为新型抗生素合成基因簇的表征提供了合适的宿主。
酿酒酵母是研究最多的模式真菌微生物,其遗传背景明晰、抗菌类次级代谢产物种类少、与外源目的基因竞争表达性小、同源重组能力强,十分有利于抗生素的研发和生产[69]。Awan等[69]通过遗传改造,在酿酒酵母中实现了青霉素的合成。Brady团队通过对酿酒酵母进行标准控制元件(如启动子)的工程改造,利用酿酒酵母实现了“沉默”抗生素合成基因簇的表达及新型抗生素的合成[70-71]。其他的酵母和丝状真菌也被尝试用于抗生素合成基因簇的表达,但研究进展相对较少[82]。
抗生素合成过程受代谢宿主微生物的严格调控[83]。利用最新的合成生物学技术,可以对细菌和真菌表达的抗生素合成基因簇进行精准调控,提高目标抗生素的产量,为大规模的临床应用提供基础。伊利诺伊大学的Wang等[72]组装了转录因子诱导物,用在链霉菌中,精确控制抑制子从“沉默”基因中脱离,进而成功“唤醒”8个大型抗生素合成基因簇,为链霉菌中天然化合物的发现及生物合成提供了新的思路。Akhter等[84]利用压力介导技术激活了海洋链霉菌菌株NA-ZhouS1中“沉默”的抗生素合成基因簇,从而分离得到新的环霉素抗生素。Lim等[73]使用CRISPR/Cas9技术将基因转录激活子——kasO*启动子插入玫瑰孢链霉菌(Streptomycesroseosporus)基因组中,激活了原本“沉默”的抗生素合成基因簇,进而发现了强效抗生素——极光霉素(Auroramycin)。CATCH和LLHR等技术可直接从放线菌染色体DNA中捕获大型抗生素合成基因簇,加快了新型抗生素异源生产的进程[75, 85]。Chen等[86]通过对链霉菌中的2个调控基因进行过表达,提高了恩拉霉素(Enramycin)的产量。张立新团队的Wang等[74]发现,链霉菌中的甘油三酯(TAG)水平与抗生素的合成密切相关,以此构建了TAG动态降解工程策略,通过精准动态调控链霉菌内甘油三酯水平,实现了多种聚酮化合物在链霉菌中的高效合成。此外,精准调控解脂耶氏酵母(Yarrowialipolytica)中TAG的降解,也能够有效提高解脂耶氏酵母生产聚酮类代谢产物的能力[87]。随着基因编辑、微生物细胞工厂构建和优化等合成生物学技术的发展,在链霉菌和酿酒酵母等宿主中异源表达抗生素合成基因簇,并通过设计—构建—测试—学习的循环对产物的产量、速率、得率等进行优化,有望使新型抗生素的规模化合成成为可能[70-71](图3)。
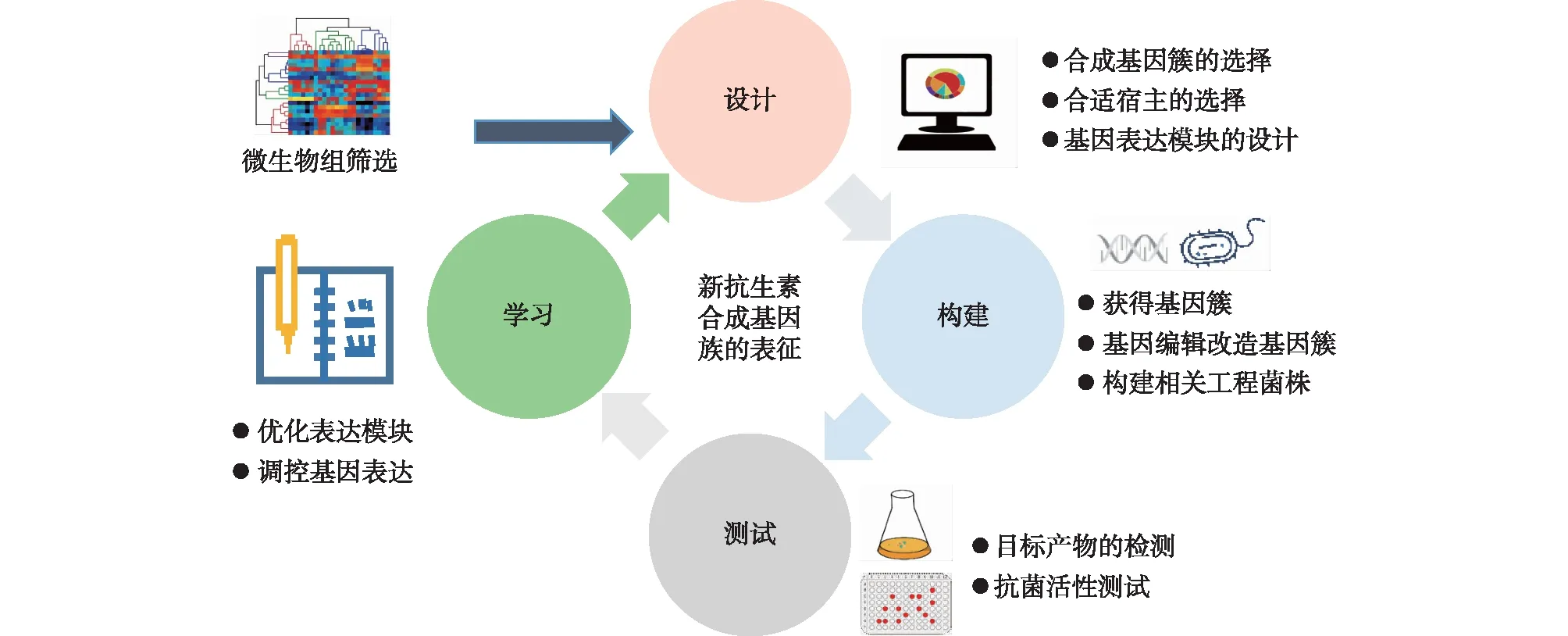
图3 合成生物学设计—构建—测试—学习循环方法表征抗生素合成基因簇
4 结论与展望
传统的依赖于可培养微生物发酵的方法已经难以发现新型抗生素。宏基因组等微生物组学技术提供了在大量未培养环境微生物中发掘新型抗生素的新策略。我们可以充分利用已有的微生物组学技术手段对环境和人体微生物群落进行解析,结合微生物代谢特征,运用培养组学技术分离未知微生物,为拓展传统可培养微生物发掘新型抗生素提供可能。同时,针对多种人体和环境微生物群落,开发高效的生物信息学和人工智能挖掘方法,筛选出合适的抗生素合成基因簇并表征,建立天然产物合成基因元件库。运用合成生物学方法和理念,表达和调控相关抗生素合成基因或者利用基因编辑手段“唤醒”沉默基因;在此基础上,利用功能基因元件组装新的抗生素合成基因簇,在链霉菌、酿酒酵母等模式宿主中表达合成相关新型抗生素,并进行抗菌活性和安全性评价,有望获得具有临床使用价值的新型抗生素。
