应用剩余杂合体衍生的近等基因系定位水稻株高QTL
2022-04-15罗先富刘文强潘孝武刘三雄刘利成阳标仁盛新年李小湘
罗先富,刘文强,潘孝武,董 铮,刘三雄,刘利成,阳标仁,盛新年,李小湘
(湖南省农业科学院水稻研究所/农业部长江中下游籼稻遗传育种重点实验室,长沙 410125)
0 引言
水稻是世界上主要的粮食作物之一,株高是影响水稻产量的重要农艺性状。根据高度不同,一般将水稻分为高秆、半矮秆和矮秆。20世纪50年代,被誉为第一次“绿色革命”的半矮秆基因sd1的成功应用使水稻单产提高了20% ~30%。随后在水稻单产实现第二次飞跃的三系杂交稻育种中,杂交稻的亲本也多采用半矮化材料。当前,株型的改良是提高水稻产量的一条重要途径,而株高是水稻株型改良的重要因子。此外,目前生产上应用的半矮秆品种基本都携带隐性基因sd1,单一基因的广泛应用存在遗传多样性丧失的潜在风险。因此,开展株高基因的定位和克隆,应用分子标记辅助选择培育适宜高度的水稻品种,具有十分重要的意义。
水稻株高的遗传主要由微效多基因控制的数量性状及主基因控制的质量性状。经典遗传学将控制株高的基因作为一个整体进行研究。林鸿宣等[1]发现粳稻‘雪禾矮早’携带有与sd-1不等位的隐性矮秆基因sd-s(t)。顾铭洪等[2]从‘桂阳矮1号’中发现了与sd-1不等位的隐性矮秆基因sd-g。随着分子生物学的发展,许多研究者利用分子标记鉴定了一系列的控制株高基因,目前已有70多个矮秆和半矮秆基因被定位(www.gramene.org)。罗炬等[3]利用重组自交系群体在水稻6条染色体上定位到控制株高的QTL,其中qPH2和qPH3在不同环境下均能检测到,并进一步精细定位qPH3在204 Kb区间。赵明芳等[4]利用构建的水稻染色体片段代换系群体共定位到3个株高相关QTL,分布在第4和第6染色体上。
近些年,矮秆和半矮秆相关基因相继被克隆。2002年,3个研究小组分别图位克隆了半矮秆“绿色革命基因”sd1,该基因编码由389个氨基酸组成的GA20氧化酶,参与赤霉素的生物合成。SD1突变导致突变体植株内源赤霉素含量降低,植株基部节间变短[5-7]。矮秆基因d1由于碱基缺失突变丧失了编码GTP结合蛋白功能,从而导致赤霉素信号传导受阻,水稻发生矮化[8]。另一个矮秆基因d61的突变导致油菜素内酯及其受体的合成受阻,造成水稻矮化[9]。D11编码BR生物合成过程的一种P450关键酶,它的突变导致油菜素内酯合成受阻,上部第二节间不伸长,植株矮化[10]。D27编码一个由278个氨基酸组成的含铁蛋白,d27突变体中第4外显子发生4 bp缺失,造成株高矮化,分蘖数增多[11]。DLT编码一个由617氨基酸组成的植物特有的GRAS家族蛋白,DLT参与BR信号传导,同时还参与BR合成的反馈抑制。DLT突变导致水稻矮化,分蘖变少[12]。水稻矮秆突变体brd1与OsBR6ox共分离,OsBR6ox编码BR-6氧化酶。突变体brd1茎秆严重矮化,株高10 ~30 cm[13]。SLR1编码产物是GA信号传导的中间调节因子,属于GRAS基因超家族,SLR1调控水稻的分蘖和株高[14]。GID1包含2个外显子,编码一条由354氨基酸组成的多肽。gid1突变体株高严重矮化、不育[15]。GID2编码一个SCF E3复合体的一个F-box亚基,介导水稻中GA的信号传导。gid2突变体严重矮化、叶片变宽、不育[16]。
尽管一些株高相关的QTL已被精细定位和克隆,但大部分QTL仍处于精度较低的初定位,所检测到的QTL大多界定在基因组的一个较大区间,仍需要进一步验证和缩小区间。此外,克隆的大多数株高基因都是属于矮秆和半矮秆基因,高秆基因的精细定位和克隆少有报道。剩余杂合体指的是从高代群体中筛选到的在目标区间表现杂合、其他区间基本纯合的单株。它相当于近等基因系材料配对杂交产生的F1材料,自交后获得的群体在目标区间呈现分离、其他区间保持纯合。利用剩余杂合体的特点,本研究从‘Katy’/‘湘743’的高代自交群体中筛选出一个高秆的剩余杂合体RHL1030,以RHL1030自交衍生群体为实验材料,开展株高QTL定位。
1 材料与方法
1.1 群体构建及标记检测
群体构建如图1所示,‘Katy’和‘湘743’杂交后代通过单粒传到F10代,经标记检测,筛选出一个在第1染色体RM11383-RM1198区间杂合(约12 Mb)、背景基本纯合的剩余杂合体RHL1030。收获其自交种子,经目标区间标记检测,从RHL1030自交群体RHL1030P中筛选出杂合区间分别为RM3411-RM11782、RM6703-RM1198的两个单株RHL1030P-13和RHL1030P-58,所覆盖的物理图谱位置分别为31.31-34.17 Mb 和 34.51-37.6 Mb(www.gramene.org),收获其自交种子种植,获得2个F2群体RHL1030P-13P和RHL1030P-58P。经SSR标记检测从RHL1030P-58P群体中筛选4个单株2-5,10-7,13-1和5-4。自交这4个单株,获得4个对应的F2群体XK2-5、XK10-7、XK13-1和XK5-4。经标记检测,分别从每个群体中筛选母本纯合型、父本纯合型及杂合型材料各40株。
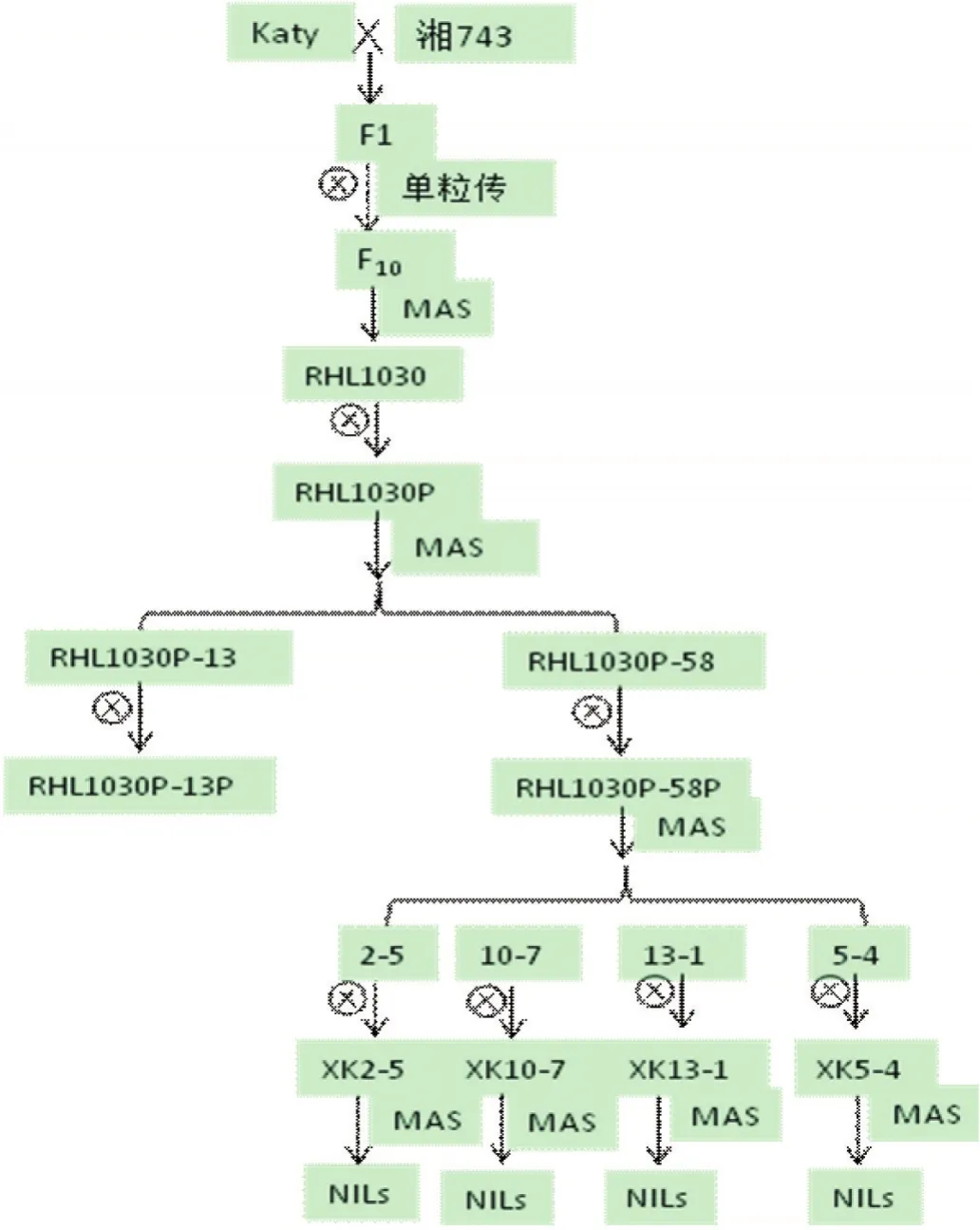
图1 群体构建示意图
本研究应用的标记均为SSR标记,采用卢等[17]的方法提取DNA,PCR扩增产物用6%的非变性聚丙烯酰胺检测。
1.2 性状考察
2019年夏在湖南农科院水稻研究所长沙实验基地种植132个单株组成的RHL1030P群体。2019—2020年在海南三亚基地种植RHL1030P-13P群体和RHL1030P-58P群体,分别包含191和151个单株。2020年夏在湖南农科院水稻研究所长沙实验基地种植XK2-5群体192株、XK10-7群体154株、XK13-1群体192株和XK4-3群体192株。株行距20×20 cm,正常田间管理。成熟后,记载群体株高。
1.3 数据分析
利用MAPMAKER(EXP3.0b)[18]作图软件,构建RHL1030P、RHL1030P-13P和RHL1030P-58P群体的区域连锁图谱,用Kosambi函数将重组率转化成遗传距离(cM)。应用 Windows QTL Cartographer 2.5[19]检测RHL1030P、RHL1030P-13P和RHL1030P-58P群体株高QTL。用SPSS对株高性状进行方差分析。
2 结果与分析
2.1 株高QTL定位
应用RHL1030P群体分离区间的7个SSR标记(RM11383、RM3411、RM297、RM212、RM6703、RM5389和RM1198)进行连锁分析,构建了第1染色体目标区间的连锁图谱,跨越25.1 cM。该群体QTL分析显示,检测到一个株高QTL,LOD值8.8,贡献率55.56%(图2)。表现加性效应为主,增效等位基因来自于父本‘湘743’。
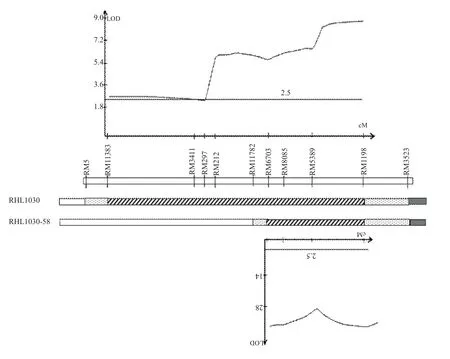
图2 利用RHL1030P群体(上)和RHL1030P-58P群体(下)进行株高QTL分析
为了进一步验证和分解目标区间,增加在‘Katy’和‘湘743’间呈现多态性的SSR标记RM11782。利用该标记从RHL1030P群体中筛选杂合区间分别为RM3411-RM11782和RM6703-RM1198的两个单株RHL1030P-13和RHL1030P-58,进一步自交发展了两个对应分离群体RHL1030P-13P和RHL1030P-58P。应用这两个群体分别进行株高QTL分析。结果显示RHL1030P-13P群体没有检测到有显著差异的QTL。RHL1030P-58P群体检测到一个显著差异的QTL,LOD值33.8,贡献率46.1%,增效等位基因来自于‘湘743’(图2)。
2.2 以染色体片段代换系效应差异为基础的QTL定位
为了进一步分解RM6703-RM1198区间,筛选并添加了一个多态性SSR标记RM8085,利用该标记从RHL1030P-58P群体中筛选到RM6703-RM8085区间杂合的3个单株2-5、10-7、13-1及RM5389-RM1198区间杂合的单株5-4。从这4个单株自交一代的群体中分别筛选分离区间为母本纯合型、父本纯合型及杂合型的单株各40株,组成4套3种不同基因型材料。通过对每一套不同基因型材料表型性状进行显著性分析,判断杂合区间是否存在显著QTL效应。
方差分析结果表明,杂合区间RM6703-RM8085分离的3个群体(XK2-5、XK10-7、XK13-1)的3种不同基因型间存在显著性差异,表明该区间存在控制株高QTL。3个群体 XK2-5、XK10-7、XK13-1检测到的QTL增效等位基因均来自于父本‘湘743’,且以加性效应为主,贡献率分别为63.1%、69.2%和74.1%。杂合区间RM5398-RM1198分离的XK5-4群体3种不同基因型间没有显著性差异。结果表明株高QTL位于RM6703-RM8085区间分离的群体。
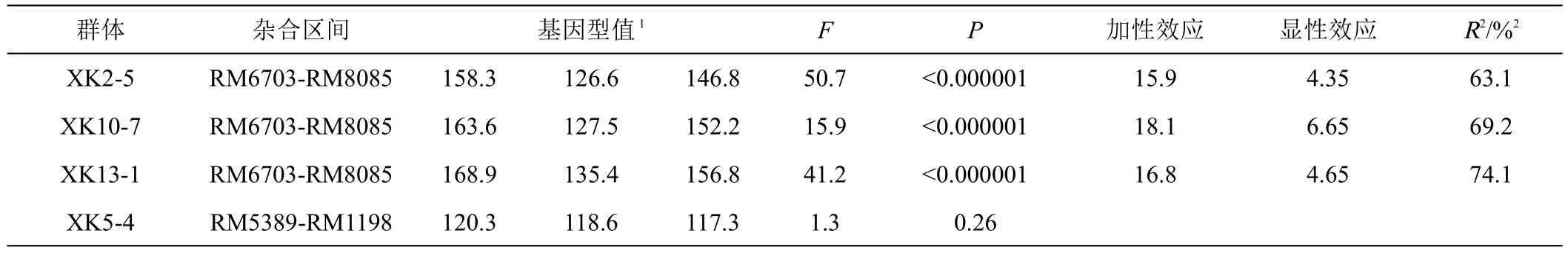
表1 2个区间在同质遗传背景下对株高性状的作用
3 讨论与结论
QTL分析在水稻重要农艺性状上已经有较多报道。但在初级定位群体中,由于遗传背景复杂,很难将QTL定位到一个精准的区间。针对初定位QTL区间,利用分子标记选择重组染色体片段代换系,在同质遗传背景下分析其性状表型差异,将目标QTL界定于更狭小的区间,是复杂性状QTL精细定位的常用方法。本研究在前期发现剩余杂合体衍生群体(RHL1030P)有株高分离的情况下进行株高QTL分析,结果在RM11383-RM1198区间鉴定到一个控制株高QTL。进而从该群体筛选不同重组单株进一步发展群体,在RM6703-RM1198区间依然能够检测到该QTL,加性效应与初定位结果一致,且进一步缩小了目标区间。同理,在RM6703-RM1198区间分离群体中筛选不同重组单株自交,通过方差分析将控制株高QTL界定在RM11782-RM5389区间。通过gramene网站查询发现,在该区间并没有控制株高基因的报道,表明本研究定位的株高相关QTL是一个新位点。
QTL定位的精细程度部分取决于多态性标记的数量。本研究证实RM6703-RM8085(34.51M ~34.86 Mb)区间分离群体包含株高QTL。由于考虑到标记两侧的重组区域,目标QTL实际界定于RM11782-RM5389区间,覆盖的物理图谱位置为34.17M ~35.73Mb。相较于RM6703-RM8085(34.51M ~34.86 Mb)区分离区间,株高QTL区间仍然较大。笔者筛选了RM8085-RM5389区间所有的SSR标记和In/Del标记,拟开展更精细的定位,遗憾的是没有筛选到多态性标记。下一步拟开发其他标记如Caps标记等进一步更精细定位株高QTL。
QTL定位受环境影响很大,相同的群体在不同的环境可能检测到不同QTL。本研究初定位群体RHL1030P种植在长沙,验证群体RHL1030P-58P群体种植在海南,均能检测到目标株高QTL,且贡献率较高,表明该QTL是一个稳定表达的主效位点。此外,在筛选剩余杂合体时,是从自交F10代开始,理论上基因组背景绝大部分纯合。因此,没有考虑目标区间外的基因组背景。XK2-5、XK10-7、XK13-1群体是在前期F10基础上继续自交两代后形成,但这3个群体在株高上仍表现明显的差异。这些结果表明尽管剩余杂合体是高代自交系,但遗传背景可能仍然存在分离。值得一提的是在XK2-5、XK10-7、XK13-1群体中,除了株高分离外,抽穗期也明显发生了分离,表明该杂合区间存在控制抽穗期相关基因。前期有报道Ghd7控制株高的同时也影响抽穗期[20],本研究定位的株高QTL是有两个控制不同性状的基因连锁亦或是一因多效还需对区间进一步分解。
本研究利用剩余杂合体自交衍生的群体定位到一个新的控制株高QTL,表明从高代群体中挑选剩余杂合体进行QTL精细定位是可行的。另一方面,鉴定的株高QTL为改良水稻株型提供更多选择。
