MAGEA6在胃癌中的表达及其意义
2022-04-01汤月良邓冠群
汤月良,邓冠群
广州市增城区人民医院普通外科,广州 511300
胃癌是消化道常见的恶性肿瘤,目前已成为全球癌症相关死亡的第二大原因。统计显示,我国每年新增胃癌患者约60万例[1]。近年来,随着内镜技术、外科手术及放化疗水平的提高,胃癌诊治效果已有一定改善,但总体生存率仍较低。由于大部分胃癌患者早期临床症状不典型,多数患者首次确诊时已是晚期,而早期确诊的胃癌经治疗,5年生存率可达90%甚至更高[2]。因此,寻找特异性强、灵敏度高的生物标志物和有效的治疗靶点,对提高胃癌的早期诊断率和生存率具有重要意义[3-4]。黑色素瘤抗原基因(melanoma antigen gene,MAGE)属于肿瘤相关抗原家族,是van der Bruggen等[5]在黑色素瘤患者的肿瘤相关抗原中发现的。MAGE家族A(MAGEA)由12个基因MAGEA1-12组成,其中MAGEA6近年来被发现与多种消化道肿瘤关系密切。据报道,MAGEA6在食管癌和肝癌组织中的表达明显高于癌旁组织,且高表达MAGEA6的食管癌和肝癌患者生存率较低[6-7]。有研究发现,MAGEA6可通过抑制肿瘤细胞自噬而促进胰腺癌的发生和发展[8],但其在胃癌组织中的表达情况、临床意义及生物学作用鲜见报道。本研究分析了胃癌组织中MAGEA6的表达情况及其与胃癌患者预后和临床病理特征的关系,并探索了其在胃癌细胞中的生物学作用。
1 资料与方法
1.1 主要试剂与仪器 BGC-823胃癌细胞系购自中科院上海细胞库;胎牛血清(fetal bovine serum,FBS)、RPMI 1640细胞培养基、青链霉素双抗、胰酶、脂质体LipofectamineTM2000转染试剂盒购自美国Thermo Fisher Scientific公司;CCK-8试剂盒、Annexin V-PE凋亡试剂盒、RIPA裂解液、BCA蛋白浓度测定试剂盒、ECL发光试剂盒购自上海碧云天生物技术有限公司;mRFP-GFP-LC3-Ⅱ腺病毒购自上海汉恒生物科技有限公司;MAGEA6、B淋巴细胞瘤-2基因(B-cell lymphoma-2,Bcl-2)、Bcl-2相关X蛋白(Bcl-2 associated X protein,Bax)、微管相关蛋白轻链3-Ⅱ(microtubule-associated protein 1 light chain 3-Ⅱ,LC3-Ⅱ)、p62蛋白(sequestosome 1,p62)、自噬相关基因5(autophagy related gene 5,Atg-5)蛋白、酵母ATG6同源物(autophagy related gene 6,Beclin 1)、蛋白激酶B(protein kinase B,Akt)、磷酸化蛋白激酶B(phosphorated protein kinase B,p-Akt)、哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)、磷酸化哺乳动物雷帕霉素靶蛋白(phosphorated mammalian target of rapamycin,p-mTOR)和甘油醛-3-磷酸脱氢酶( glyceraldehyde-3-phosphate dehydrogenase,GAPDH)单克隆抗体购自英国Abcam公司;免疫组化试剂盒购自福州迈新生物技术开发有限公司。微光分光光度计购自美国Merinton公司;Mini-Proten Tetra System电泳系统和ChemiDoc XRS+System凝胶成像仪购自美国Bio-Rad公司;ACCURI C6流式细胞仪购自美国BD公司;Tecan Infinite Pro全波长多功能酶标仪购自中国Tecan公司;倒置显微镜购自日本Olympus公司;激光共聚焦显微镜购自美国Zeiss公司;透射电镜购自荷兰Philips公司。
1.2 胃癌组织芯片 胃癌组织芯片样本90例由本院样本库提供,为2009年12月-2010年6月经手术切除的胃癌组织和癌旁组织,其中男68例,女22例。纳入标准:(1)病理诊断为胃部腺癌、印戒细胞癌和黏液癌;(2)既往未进行放疗和化疗;(3)行根治性胃癌切除术。排除标准:(1)胃恶性间质瘤;(2)神经内分泌癌;(3)因手术并发症死亡;(4)病历资料不完整。
1.2.1 免疫组化检测胃癌组织芯片中MAGEA6的表达情况 将芯片组织进行脱蜡、水化,H2O2阻断内源性过氧化物酶,使用EDTA/枸橼酸钠溶液进行抗原修复,加入MAGEA6(1∶200)一抗4 ℃下孵育过夜;PBS清洗3次,加入生物素标记的二抗(1∶200)室温孵育30 min;PBS清洗3次,加入链霉菌抗生物素过氧化物酶溶液室温孵育20 min;PBS清洗4次,行DAB染色,然后行苏木精染色;清洗后乙醇脱水、二甲苯透明,使用中性树胶固定封片,显微镜下观察,对每个视野进行染色强度计分和阳性细胞百分比评分。染色强度计分:无明显着色、轻微着色、中度着色和重度着色分别计为0、1、2、3分;阳性细胞百分比评分:阳性细胞百分比≤5%、5%~25%、25%~50%、50%~75%和>75%分别计为0、1、2、3、4分。最终评分=阳性细胞百分比评分+染色强度计分。最终评分≥4分为高表达,<4分为低表达。
1.2.2 MAGEA6表达与胃癌临床病理特征的关系
收集胃癌患者性别、年龄、肿瘤大小(<5 cm;≥5 cm)、肿瘤部位(胃窦、胃体、贲门)、淋巴转移(阴性/阳性)、病理分型(腺癌、印戒细胞癌、黏液癌)、TNM分期(Ⅰ、Ⅱ、Ⅲ、Ⅳ期)等一般资料。依据MAGEA6的表达情况,将胃癌患者分为MAGEA6高表达组与MAGEA6低表达组,分析MAGEA6表达与胃癌临床病理特征的关系。
1.2.3 生存分析 采用Kaplan-Meier Plotter 40(KM Plotter)在线工具(http://kmplot.com/),选择基因芯片数据集GSE62254、GSE15459、GSE29272分析胃癌患者的MAGEA6表达情况及其预后,生存曲线图、病例数、风险比(hazard ratio,HR)及log-rankP值从数据库中获取。90例胃癌患者的生存情况采用GraphPad Prism 8.0软件进行分析。
1.3 细胞分组及处理 BGC-823细胞于RPMI 1640(含10% FBS和1%青链霉素双抗)完全培养基中培养,置于37 ℃、5% CO2、饱和湿度的恒温细胞培养箱中;当细胞融合度为70%~80%时,用胰蛋白酶消化,按照1∶2的比例传代。取对数生长期细胞接种于6孔板中(2×105个/孔),当细胞融合度达到50%时,设置si-MAGEA6组与si-Ctrl组,参照LipofectamineTM2000转染说明书操作,si-MAGEA6组加入10 μlsi-MAGEA6转染,si-Ctrl组加入150 μl siRNA空载体对照转染,转染24 h后更换培养基,并进行后续实验。
1.3.1 CCK-8法检测细胞活力 将转染后的BGC-823细胞接种于96孔板中(密度为5×103个/ml),分别在转染后24、48和72 h检测细胞活性。每孔加入10 μl CCK-8试剂,37 ℃孵育4 h,使用酶标仪检测各孔450 nm处的吸光度(OD)值。
1.3.2 流式细胞术检测细胞凋亡情况 离心并收集细胞,取2×105个细胞,PBS清洗3次,加入Annexin V-FITC结合液重悬细胞,依次加入Annexin V/FITC和PI,室温避光孵育20 min,在避光条件下上机检测细胞凋亡情况。
1.3.3 Western blotting检测MAGEA6、凋亡相关蛋白、自噬相关蛋白及Akt/mTOR信号通路相关蛋白的表达 收集细胞,使用RIPA裂解液提取总蛋白。使用SDS-PAGE凝胶电泳分离蛋白,并转至PVDF膜上,使用5%脱脂奶粉封闭2 h;加入MAGEA6、Bcl-2、Bax、LC3-Ⅱ、p62、Atg-5、Beclin 1、Akt、p-Akt、mTOR、p-mTOR和GAPDH单克隆一抗(1∶1000)并于4 ℃摇床中孵育过夜;PBST洗膜3次,加入兔二抗(1∶1000)避光孵育1 h;PBST洗膜3次,使用Odyssey IR凝胶成像仪进行扫描并拍照,ImageJ软件进行灰度值分析,计算目的蛋白与相应内参条带灰度值的比值。
1.3.4 扫描电镜观察自噬小体形成情况 收集细胞,PBS清洗3次,使用戊二醛(3%)固定细胞,丙酮梯度脱水;将细胞包埋在环氧树脂中,切片,使用枸橼酸铅和醋酸铀染色,透射电镜观察细胞中自噬小体形成情况。
1.4 激光共聚焦显微镜观察细胞自噬流变化 将对数生长期BGC-823细胞接种于6孔板中(2×105个/孔),用si-MAGEA6转染24 h,然后用感染复数为300的mRFP-GFP-LC3-Ⅱ腺病毒感染24 h,收集细胞爬片,4%多聚甲醛溶液室温固定,封片,激光共聚焦显微镜下观察BGC-823细胞内自噬流的变化。
1.5 统计学处理 使用GraphPad Prism 8.0软件进行统计分析。计量数据以±s表示,符合正态分布且方差齐时,两组间比较采用Student'st检验;计数资料以率(%)表示,两组间比较采用χ2检验。使用Kaplan-Meier在线工具进行生存分析,使用log-rank法对MAGEA6高表达组和低表达组患者的生存率进行比较。P<0.05为差异有统计学意义。
2 结 果
2.1 MAGEA6在胃癌组织中的表达情况 免疫组化检测结果显示,MAGEA6主要表达于细胞质中(图1A);胃癌组织MAGEA6免疫组化评分明显高于癌旁组织[(3.77±1.50)分vs. (2.58±1.11)分,P<0.05,图1B);胃癌组织MAGEA6高表达率明显高于癌旁组织[57.8%(52/90)vs. 20.0%(18/90),χ2=27.02,P<0.001,图1C]。

图1 胃癌组织与癌旁组织中MAGEA6的表达情况Fig.1 Expression of MAGEA6 in gastric cancer and paracancerous tissues
2.2 MAGEA6表达与胃癌临床病理特征的关系MAGEA6表达水平与胃癌患者的性别、肿瘤大小、肿瘤部位、病理分型及有无淋巴转移无关(P>0.05),而与年龄、TNM分期有关(P<0.05,表1)。

表1 MAGEA6表达与胃癌患者临床病理特征的关系[例(%)]Tab.1 Relationship between the expression of MAGEA6 and the clinicopathological characteristics of gastric cancer patients[n(%)]
2.3 MAGEA6表达与胃癌预后的关系 Kaplan-M e i e r P l o t t e r 生存分析结果显示,基因芯片GSE62254、GSE29272数据集中MAGEA6高表达的胃癌患者中位生存期与MAGEA6低表达患者比较差异无统计学意义(GSE62254:20.13个月vs. 25.87个月,P=0.230;GSE29272:22.00个月vs. 25.90个月,P=0.066);GSE15459数据集中MAGEA6高表达的胃癌患者中位生存期短于MAGEA6低表达的患者(26.50个月vs. 30.40个月,P=0.021) (图2A-C)。
对本组90例胃癌患者进行生存分析,结果显示,MAGEA6高表达组患者的中位生存期短于MAGEA6低表达组,但差异无统计学意义(25.50个月vs. 64.50个月,P=0.080,图2D)。

图2 MAGEA6表达与胃癌患者预后的关系Fig.2 Relationship between MAGEA6 and prognosis of patients with gastric cancer
2.4 敲减MAGEA6表达对胃癌细胞增殖和凋亡的影响 CCK-8法检测结果显示,与si-Ctrl组比较(48 h:1.33±0.10;72 h:1.79±0.14),si-MAGEA6组胃癌细胞在转染48 h(0.80±0.07)和72 h(1.15±0.09)的OD450明显降低(P<0.05,图3A)。
流式细胞术检测结果显示,与si-Ctrl组比较,si-MAGEA6组细胞凋亡率明显增高(14.97%±0.86%vs. 4.63%±0.55%,P<0.05,图3B)。

图3 敲减MAGEA6表达对胃癌细胞增殖和凋亡的影响Fig.3 Effect of knockdown of MAGEA6 expression on proliferation and apoptosis of gastric cancer cells
Western blotting检测结果显示,si-MAGEA6组MAGEA6、Bcl-2蛋白表达水平低于si-Ctrl组(MAGEA6:0.33±0.04vs. 0.69±0.06,P<0.05;Bcl-2:0.37±0.04vs. 0.54±0.06,P<0.05),Bax蛋白表达水平高于si-Ctrl组(0.62±0.05vs. 0.28±0.04,P<0.05,图4)。

图4 敲减MAGEA6表达对胃癌细胞中MAGEA6、Bcl-2和Bax蛋白表达的影响(Western blotting)Fig.4 Effect of knockdown of MAGEA6 expression on the expression of MAGEA6, Bcl-2 and Bax proteins in gastric cancer cells(Western blotting)
2.5 敲减MAGEA6表达对胃癌细胞自噬的影响
Western blotting检测结果显示,si-MAGEA6组自噬相关蛋白Atg-5、Beclin 1表达水平以及LC3-Ⅱ/LC3-Ⅰ高于si-Ctrl组(Atg-5:0.41±0.08vs. 0.19±0.04,P<0.05;Beclin 1:0.24±0.04vs.0.12±0.03,P<0.05;LC3-Ⅱ/LC3-Ⅰ:0.61±0.08vs. 0.33±0.07,P<0.05),p62蛋白表达水平低于si-Ctrl组(0.24±0.05vs. 0.43±0.05,P<0.05,图5)。
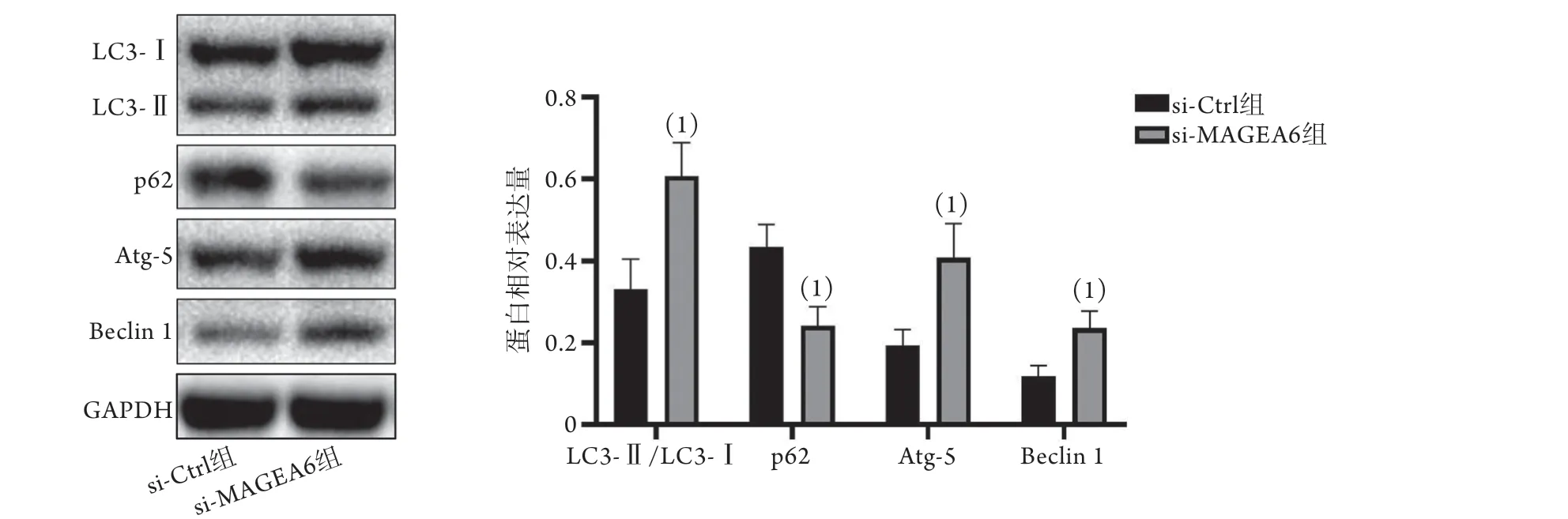
图5 敲减MAGEA6表达对自噬相关蛋白表达的影响(Western blotting)Fig.5 Effect of knockdown of MAGEA6 expression on the expression of autophagy-related proteins (Western blotting)
为进一步明确敲减MAGEA6表达对胃癌细胞自噬的影响,利用电镜和激光共聚焦显微镜观察胃癌细胞内自噬小体形成情况及自噬流的变化。电镜观察显示,与si-Ctrl组比较,si-MAGEA6组胃癌细胞胞质内自噬小体明显增多(图6);激光共聚焦显微镜观察显示,敲减MAGEA6表达可促进胃癌细胞内自噬小体与溶酶体形成自噬溶酶体,提示胃癌细胞自噬增强(图7)。

图6 电镜观察敲减MAGEA6表达对胃癌细胞自噬小体形成的影响Fig.6 Electron microscopic observation the effect of knockdown of MAGEA6 expression on the autophagosome formation in gastric cancer cells
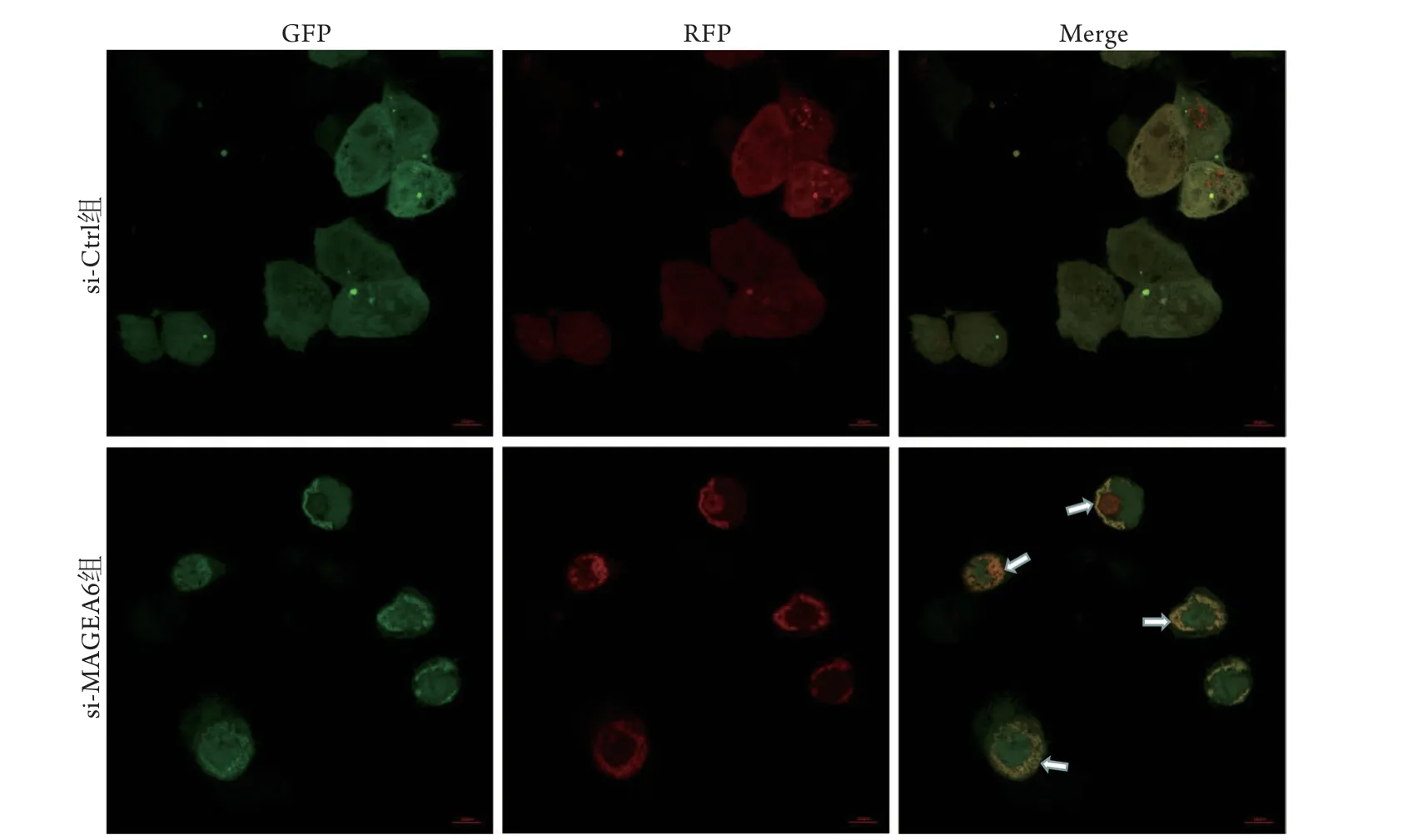
图7 激光共聚焦显微镜观察敲减MAGEA6表达后胃癌细胞自噬流的变化Fig.7 Laser confocal microscope observation the changes of autophagic flux after knockdown MAGEA6 expression of gastric cancer cells
2.6 敲减MAGEA6表达对胃癌细胞Akt/mTOR信号通路的影响 Western blotting检测结果显示,si-Ctrl组和si-MAGEA6组Akt、mTOR总蛋白表达水平差异无统计学意义(P>0.05);与si-Ctrl组相比,si-MAGEA6组p-Akt、p-mTOR蛋白表达水平明显降低[p-Akt:0.47±0.07vs. 0.90±0.07,P<0.05;p-mTOR:0.34±0.04vs. 0.58±0.04,P<0.05,图8]。

图8 敲减MAGEA6表达对胃癌细胞Akt、mTOR、p-Akt、p-mTOR蛋白表达的影响(Western blotting)Fig.8 Effect of knockdown of MAGEA6 expression on Akt, mTOR, p-Akt, and p-mTOR protein expression of gastric cancer cells(Western blotting)
3 讨 论
胃癌为消化道的恶性肿瘤,其发病率较高,我国胃癌患者约占全球胃癌总数的50%,严重威胁着人民群众的健康。我国东部地区的胃癌发病率高于西部地区,吸烟、亚硝酸盐和酒精摄入均能增加胃癌的患病风险,且大部分患者在发现时已处于Ⅲ期或Ⅳ期[9-11]。胃癌晚期进行手术治疗、放疗和化疗已很难提高生存率,且癌细胞容易对化疗药物产生耐药性[11-12]。因此,阐明胃癌发生和发展的分子机制对于研发有效的治疗药物具有重要意义。MAGE家族首先在黑色素瘤患者中发现,是黑色素瘤相关抗原[5]。MAGE超家族包含60多个基因,具有A、B和C三个亚家族,在进化中高度保守[13-15]。其中MAGEA具有肿瘤特异性,可针对其研制肿瘤疫苗[16]。研究发现,MAGEA对肿瘤的发生发展和药物抵抗具有重要作用[17]。MAGEA高表达与多种肿瘤预后不良有关[18-19]。本研究发现,MAGEA6在胃癌组织中高表达,且高表达MAGEA6的胃癌患者总体生存率低于低表达的患者。MAGEA6高表达与年龄及临床分期有关,提示MAGEA6可作为胃癌潜在的诊断和预后评估标志物以及治疗靶点。
研究发现,在肿瘤的发生过程中细胞凋亡的自我调节被打破。Bcl-2和Bax是调节细胞凋亡的关键蛋白,Bcl-2/Bax比值增高可抑制细胞凋亡,比值降低则可促进细胞发生凋亡[20]。细胞自噬是维持细胞内环境稳态的重要自我调节机制,对于细胞行为学和功能影响较大,与肿瘤的发生和发展关系密切[21]。调控自噬的分子信号通路十分复杂,当自噬发生时,胞质型LC3(LC3-Ⅰ)可酶解掉一小段多肽转变为膜型(LC3-Ⅱ),在膜内外表达的LC3-Ⅱ会脱离自噬体而进入细胞质,所以LC3-Ⅱ与LC3-Ⅰ的比值可反映细胞自噬的水平[22-24]。Beclin-1是自噬小体形成的关键蛋白,可启动细胞自噬,上调Beclin-1可诱导细胞自噬的发生[25-26]。在内质网应激过程中,自噬可消化对细胞生存无意义的蛋白质来维持内质网的功能,进而减少内质网应激诱发的凋亡,而过度自噬可加速细胞自噬性细胞死亡[27]。本研究发现,敲减MAGEA6的表达可抑制胃癌细胞的增殖能力,促进胃癌细胞凋亡,抑制抗凋亡蛋白Bcl-2的表达,促进凋亡蛋白Bax的表达;此外还可促进胃癌细胞自噬。这与既往研究中MAGEA6参与调控多种癌细胞自噬水平的结果一致[8,16]。但敲减MAGEA6促进胃癌细胞凋亡是否与其增强胃癌细胞自噬有关仍需进一步探讨。
Akt/mTOR信号通路主要由PKB/Akt和mTOR两个作用分子组成。Akt在细胞存活和凋亡中起重要作用,其Ser473位点可被PDK1磷酸化,是Akt/mTOR信号通路的中心作用分子;mTOR是Akt的下游分子,是一种营养和能量状态的传感器,受多种信号的调控,可通过被Akt直接或间接磷酸化而激活[28-30]。大量研究发现,肿瘤细胞自噬与Akt/mTOR信号通路关系密切。雷公藤根皮中主要的活性天然产物雷公藤红素在脑胶质瘤细胞中可通过降低Akt/mTOR通路中p-Akt、p-mTOR的表达水平而增强细胞自噬,促进细胞凋亡[31]。3'-epi-12β-羟基脯氨酸可通过阻断肺癌细胞中Akt/mTOR通路而诱导细胞保护性自噬,进而发挥抑制肺癌细胞生长的作用[32]。本研究发现,敲减MAGEA6后细胞自噬水平明显升高,且Akt、mTOR磷酸化水平降低,提示si-MAGEA6抑制Akt/mTOR通路的磷酸化是其增强胃癌细胞自噬的潜在作用机制。
综上所述,本研究结果表明,MAGEA6在胃癌组织中呈高表达,敲减MAGEA6能够抑制胃癌细胞增殖、促进胃癌细胞凋亡,还可通过抑制Akt/mTOR通路的磷酸化而介导自噬增强。MAGEA6有望成为胃癌诊断和预后评估的特异性生物标志物以及潜在的治疗靶点。但本研究未探索MAGEA6介导的自噬与凋亡的关系,也未通过体内试验进行验证,后续研究需建立Akt/mTOR通路高表达和低表达的胃癌细胞进行机制验证,以为胃癌的诊断和治疗提供新的途径。
