干旱胁迫下CO2浓度倍增对达乌里胡枝子幼苗生长及抗氧化特性的影响
2022-03-28杨耀国牛冰洁王永新卫昭君郭媛珍王开喜朱慧森
杨耀国, 牛冰洁, 王永新*, 卫昭君, 郭媛珍, 王开喜, 马 春, 赵 祥, 朱慧森
(1.山西农业大学草业学院, 山西 太谷 030801; 2.草地生态保护与乡土草种质创新山西省重点实验室, 山西 太谷 030801)
近年来,全球工业的迅速发展和人类的活动导致大气中温室气体浓度不断升高,IPCC第五次报告预计本世纪末CO2浓度可能达到421~936 μmol·mol-1[1]。CO2浓度持续升高会引起温室效应和增加极端降水事件[2],据预测未来100年地表温度会升高2.2℃[3],温度升高会使土壤含水量下降从而导致植物干旱[4]。干旱不仅影响植物的生长速度,而且影响植物的生理响应[5],导致植物生长发育产生严重阻碍;同时会使植物体内发生膜脂过氧化,产生大量活性氧自由基,导致细胞膜结构被破坏且相对电导率会升高[6]。干旱时间的增加,会导致细胞脱水,丙二醛(Malondialdehyde,MDA)和脯氨酸(Proline,Pro)不断积累,可溶性糖(Soluble sugar,SS)和可溶性蛋白(Soluble protein,SP)含量也会持续升高[7],同时植物可通过增强超氧化物歧化酶(Superoxide dismutase,SOD)和过氧化物酶(Peroxidase,POD)来清除活性氧自由基以避免干旱对其造成伤害,从而增强抗旱性[8]。而植物生长除受水分影响外,也与CO2浓度高低有密切联系[9]。CO2是植物生长和光合作用的底物,CO2浓度的升高会促进植物的光合作用,提高水分利用率,从而影响植物的生长发育[10]。研究表明,高浓度CO2可以促进植物体内产生一系列抗氧化酶,在一定程度上激活抗氧化防御系统,抑制活性氧(Reactive oxygen species,ROS)的积累,进而提高植物遭受干旱胁迫后各功能的恢复能力[11]。牛胤全等[12]通过研究发现在干旱胁迫下高浓度CO2可以促进小麦(Triticumaestivum)叶面积增加,同时促进根部分蘖和根系生长,在一定程度上增强其幼苗的抗氧化性。肖列等[13]通过试验得出CO2浓度升高能够缓解干旱胁迫造成的不利影响,提高了白羊草(Bothriochloaischaemum)的抗旱性。
达乌里胡枝子(Lespedezadavurica)为豆科(Leguminosae)胡枝子属(Lespedeza)多年生草本植物[14],国内主要分布在河北、东北、内蒙古、华北、西北等地区[15],国外主要分布在蒙古、俄罗斯、朝鲜、日本等国。因其蛋白质和脂肪含量高,有较好的适口性,可以作为优良的牧草应用于饲料加工[16]。其根系发达且适应性强,具有耐阴、耐寒、耐干旱耐瘠薄等特点[17],在改善生态环境、改良退化草地和推进退耕还林工程中发挥着极其重要的作用[18]。现国内有关达乌里胡枝子的研究主要集中于抗旱性[19-21]、生产特性[22-24]等方面,对干旱胁迫和高CO2浓度相互作用的生理机制研究相对较少。本试验通过研究达乌里胡枝子幼苗在干旱胁迫下抗氧化酶活性的变化,探讨CO2倍增与干旱胁迫达乌里胡枝子幼苗抗氧化特性的影响,旨在为适应未来气候变化条件下达乌里胡枝子的种质资源评价提供参考依据。
1 材料与方法
1.1 试验材料
供试材料为山西农业大学草业学院自主选育的‘晋农1号’达乌里胡枝子。
1.2 试验设计
1.2.1生长条件 本试验采用盆栽方式,在山西农业大学草业学院校内试验地取土,每盆装风干土2.0 kg,每盆均匀播种50粒。将花盆放置于温室中进行发芽,温室平均温度昼/夜分别为29℃/14℃,相对湿度昼/夜分别为79%/64%,幼苗长至10 cm时进行定苗,使每盆保留30株长势相近的幼苗,随机移入两个人工气候培养箱(RCL-P1000-CO2型),每个培养箱放置32盆,共计64盆,适应1周后进行干旱胁迫。
CO2处理由2个人工气候培养箱分别控制,设定温度和湿度分别为(27±1)℃和(60±2)%,模拟自然光光照强度。
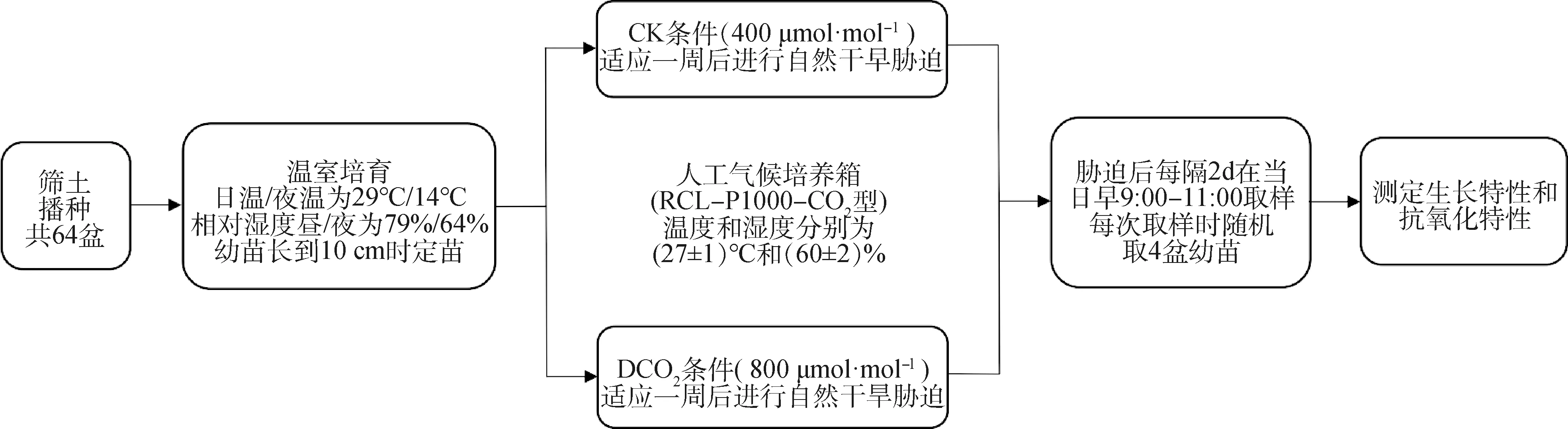
图1 试验示意图Fig.1 Experimental schematic diagram
1.2.2干旱处理和CO2浓度设定 设置两个CO2浓度:正常CO2浓度(400 μmol·mol-1,CK)和倍增CO2浓度(800 μmol·mol-1,DCO2)。通过自然消耗的方法进行干旱胁迫处理,依据D-最优性法则和预试验结果,开始观测前对所有盆栽同时进行浇透水处理(干旱胁迫第0 d),之后不再浇水使其自然干旱,干旱胁迫的第2,4,6,8,10,12,14,16 d分别在每日上午9∶00—11∶00取样,每次取样时随机取4盆幼苗进行测定。
1.3 生长特性的测定
1.3.1幼苗株高测定 每盆随机选取10株幼苗,用直尺测量株高,将全株的地上与地下部分离并清洗干净后,称量地上部所有植株鲜重,计算株高和鲜重相对生长速率。
相对生长速率=以2 d为时间间隔所测定的每盆选取的10株达乌里胡枝子指标平均值的差值/时间[25]。
1.3.2幼苗主根长测定 每盆随机选取10株植株,将地上部与地下部分离并清洗干净后,用直尺测量主根长,称量地下部所有根系鲜重,计算主根长和根鲜重相对生长速率。
1.4 抗氧化特性的测定
1.4.1相对含水量和相对电导率的测定 在每个测定日的上午11点,选取生长健壮的植株剪取三小叶的中间小叶10片,称量鲜重(W1)后放入烘箱,105℃杀青15 min后烘干至恒重(W2),计算叶片相对含水量(Leaf relative water content,LRWC)。
LRWC=(W1—W2)/W1×100%[26]
选取生长健壮的植株剪取三小叶的中间小叶5片,蒸馏水冲洗3次后剪成大小相近的碎片,采用电导仪法[26]进行相对电导率的测定。
1.4.2抗氧化酶活性和渗透调节物质的测定 采用氮蓝四唑法[27]测定SOD活性,愈创木酚比色法[27]测定POD活性,紫外分光光度计法[27]测定SP含量,苯酚法[27]测定SS含量,磺基水杨酸提取法[27]测定Pro含量,硫代巴比妥酸比色法[27]测定MDA含量。
1.5 数据分析
用Microsoft Excel 2016进行数据整理和计算,首先对数据进行正态性检验,后用SPSS 23.0进行方差分析,并采用Duncan法进行差异显著性的多重比较,采用Pearson法做相关性分析。使用Sigma Plot 12.5进行绘图。
2 结果与分析
2.1 干旱胁迫下CO2浓度倍增对达乌里胡枝子幼苗生长特性的影响
2.1.1干旱胁迫下CO2浓度倍增对达乌里胡枝子幼苗植株的影响 CK条件下,株高相对生长速率随着自然干旱处理天数增加呈下降趋势。在CO2浓度倍增条件下,达乌里胡枝子幼苗株高相对生长速率随着自然干旱处理天数的增加呈先上升后下降的趋势(图2a),在干旱胁迫至第6 d时,相对生长速率升至最高。除第14 d外,CO2浓度倍增条件下达乌里胡枝子幼苗的株高相对生长速率与CK处理差异均达到显著水平(P<0.05)。干旱胁迫至第16 d时,两个CO2浓度处理下的株高相对生长速率均达到最低,CO2浓度倍增条件下株高生长速率较CK提高93.94%。
不同CO2浓度条件下,随着自然干旱处理天数的增加,达乌里胡枝子幼苗的植株鲜重相对生长速率均呈下降的趋势(图2b)。干旱胁迫6~12 d时呈显著性下降趋势,胁迫至12~16 d时下降趋势变缓。CO2倍增条件下达乌里胡枝子幼苗的植株鲜重相对生长速率均高于CK,在干旱胁迫4~6 d时CO2浓度倍增处理显著高于CK处理(P<0.05),干旱胁迫至第16 d时CO2倍增处理比CK提高1.04%。

图2 干旱胁迫下CO2浓度倍增对达乌里胡枝子幼苗植株的影响Fig.2 Effects of doubling CO2 concentration on Lespedeza davurica seedling under drought stress
2.1.2干旱胁迫下CO2浓度倍增对达乌里胡枝子幼苗主根长的影响 不同CO2浓度条件下达乌里胡枝子幼苗的主根长相对生长速率均随着自然干旱处理天数的增加呈先增加后降低的趋势(图3a)。在CO2倍增条件下,干旱胁迫4~14 d时,主根长相对生长速率持续上升,干旱胁迫至第14 d时,主根长相对生长速率最大,后随干旱胁迫天数增加呈下降趋势;CK条件下,在干旱胁迫4~12 d时,主根长相对生长速率持续上升,在干旱胁迫至第12 d时主根长相对生长速率最大,后随干旱胁迫天数增加呈下降趋势。CO2倍增条件下达乌里胡枝子幼苗的主根长相对生长速率峰值出现时间晚于CK,且与CK差异显著(P<0.05),CO2倍增处理下干旱胁迫至第14 d时主根长相对生长速率较CK处理下干旱胁迫至第12 d时显著提升61.88%(P<0.05),干旱胁迫至第16 d时,CO2倍增处理较CK显著提升91.73%(P<0.05)。
不同CO2浓度条件下达乌里胡枝子幼苗的根系鲜重相对生长速率均随着自然干旱处理天数的增加呈缓慢上升后下降的趋势,且CO2倍增条件下根系鲜重相对生长速率峰值出现比CK晚(图3b)。CO2倍增条件下,干旱胁迫至4~12 d时,达乌里胡枝子幼苗的根系鲜重相对生长速率缓慢上升,干旱胁迫至14 d时,根系鲜重相对生长速率达最大值,较CK处理下干旱胁迫最大值(第12 d时)显著提升37.15%(P<0.05),干旱胁迫至14~16 d时,根系鲜重相对生长速率呈显著下降趋势(P<0.05)。CK条件下,干旱胁迫至4~10 d时,达乌里胡枝子幼苗的根系鲜重相对生长速率缓慢上升,干旱胁迫至10~12 d时根系鲜重相对生长速率呈显著上升趋势(P<0.05),干旱胁迫至12~16 d时,根系鲜重相对生长速率呈显著下降趋势(P<0.05)。干旱胁迫至第16 d时,CO2倍增条件下根系鲜重相对生长速率比CK显著提高246.38%(P<0.05)。
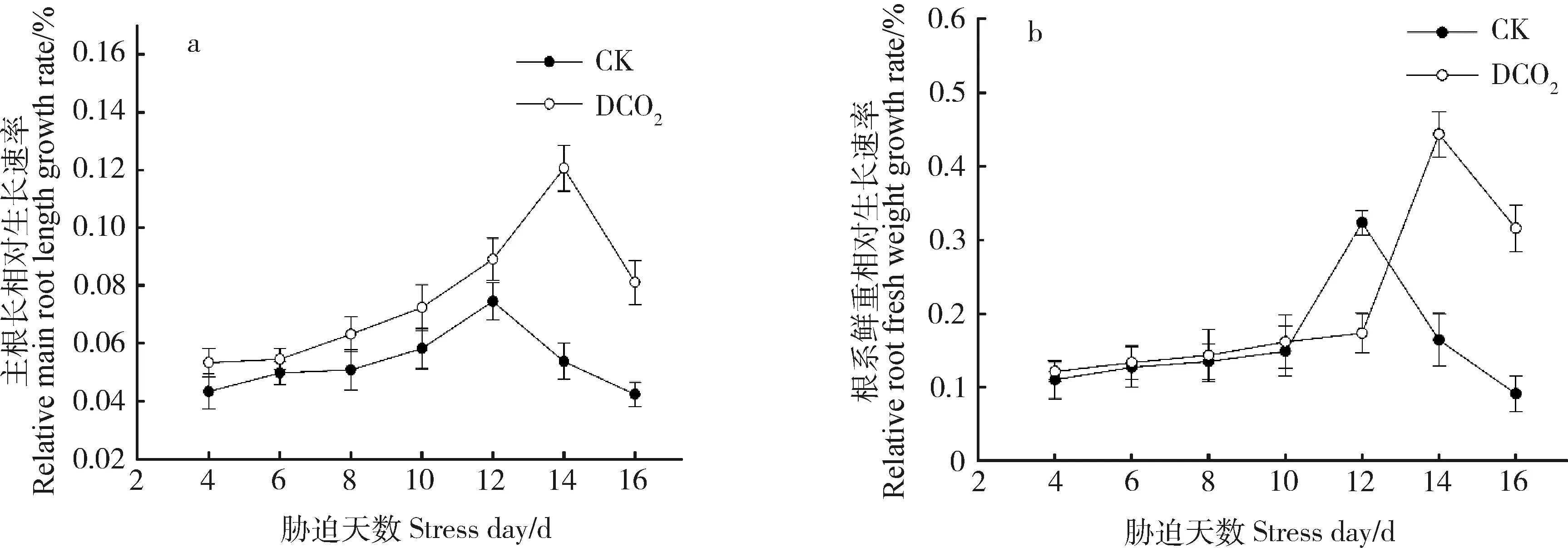
图3 CO2倍增和干旱胁迫对达乌里胡枝子幼苗主根的影响Fig.3 Effects of CO2 doubling and drought stress on the taproot of Lespedeza davurica seedlings
2.2 干旱胁迫下CO2浓度倍增对达乌里胡枝子幼苗抗氧化特性的影响
2.2.1干旱胁迫下CO2浓度倍增对达乌里胡枝子幼苗抗氧化酶的影响 图4a为CO2倍增和干旱胁迫对达乌里胡枝子幼苗SOD活性的影响。如图所示,随着自然干旱处理天数的增加,两种CO2浓度条件下SOD活性均呈先升高后降低的趋势,CO2倍增条件下的SOD活性始终高于CK,且随着自然干旱处理天数的延长两者差异逐渐变大。干旱胁迫和CO2浓度倍增对SOD活性的影响具有显著的交互作用,其中CO2浓度倍增条件下达乌里胡枝子SOD活性峰值出现时间晚于CK,且与CK差异显著(P<0.05)。
图4b为CO2倍增和干旱胁迫对达乌里胡枝子幼苗POD活性的影响。如图所示,随着自然干旱处理天数的延长,两种CO2浓度条件下POD活性均呈先升高后降低的趋势。CO2浓度倍增条件下,干旱胁迫至第8 d时,POD活性与干旱胁迫第2 d时无显著差异,干旱胁迫至第12 d时POD活性达最大值,较干旱胁迫至第2 d时增加了24.3%,干旱胁迫和CO2浓度倍增对POD的影响具有显著的交互作用,在干旱胁迫的12~14 d时CO2倍增下POD活性显著高于CK。
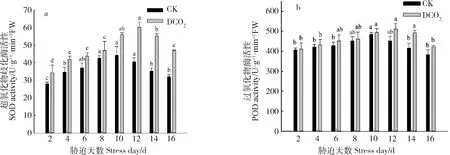
图4 CO2倍增和干旱胁迫对达乌里胡枝子幼苗抗氧化酶活性的影响Fig.4 Effect of doubled CO2 on antioxidant enzyme activity of Lespedeza davurica seedling under drought stress注:不同小写字母表示相同CO2浓度不同干旱胁迫天数之间差异显著(P<0.05),DCO2为CO2浓度倍增条件,下同Note:Different lowercase letters indicate significant difference between different drought stress days with the same CO2 concentration at the 0.05 level,DCO2 is the doubling condition of CO2 concentration,the same as below
干旱胁迫至第16 d时,CO2倍增条件下SOD活性较CK提高了46.28%(P<0.05),POD活性较CK提高了10.18%。
2.2.2干旱胁迫下CO2浓度倍增对达乌里胡枝子幼苗渗透调节物质和LRWC的影响 图5a为CO2倍增和干旱胁迫对达乌里胡枝子幼苗SP含量的影响。如图所示,CK条件下,干旱胁迫至第4 d时,SP含量达到最大值,干旱胁迫至第14 d时出现最小值;在CO2倍增条件下,干旱胁迫第6 d时SP含量最低,干旱胁迫至第10 d时SP含量达到最大值,后随干旱胁迫天数的增加SP含量开始降低。
图5b为CO2倍增和干旱胁迫对达乌里胡枝子幼苗SS含量的影响。如图所示,在干旱胁迫至2~4 d时,两种CO2浓度条件下达乌里胡枝子幼苗SS含量均上升,干旱胁迫4~8 d时均呈下降趋势,干旱胁迫10~16 d时均呈上升趋势,干旱胁迫至第16 d时两种CO2浓度条件下的SS含量差异显著(P<0.05)且均达到最大值。
图5c为CO2倍增和干旱胁迫对达乌里胡枝子幼苗Pro含量的影响。如图所示,随着自然干旱处理天数的延长,两种CO2浓度条件下达乌里胡枝子幼苗的Pro含量逐渐增大,CO2倍增条件下Pro含量均高于CK,干旱胁迫至第16 d时,CO2倍增条件下Pro含量较CK显著提高41.37%(P<0.05)。CO2浓度倍增条件下,干旱胁迫至第14 d时,达乌里胡枝子幼苗的Pro含量变化出现显著差异(P<0.05);两种CO2浓度条件下均在干旱胁迫至第16 d时达到最大值,CK条件下16 d时Pro含量较干旱胁迫第2 d时上升169.2%。
图5d为CO2倍增和干旱胁迫对达乌里胡枝子幼苗LRWC的影响。如图所示,两种CO2浓度条件下,达乌里胡枝子幼苗的LRWC均随着自然干旱处理天数的延长而减小。CO2浓度倍增条件下的LRWC均高于CK,且随着自然干旱处理天数的延长差异逐渐变大,干旱胁迫至第16 d时,CO2倍增条件下LRWC较CK提高48.80%(P<0.05)。CK条件下干旱胁迫超过第10 d时LRWC急速减少,而CO2倍增条件下LRWC缓慢减少;干旱胁迫至第16 d与干旱胁迫至第2 d时LRWC差异显著(P<0.05),CK和CO2倍增条件下LRWC分别减少了46.2%和20.2%。

图5 CO2倍增和干旱胁迫对达乌里胡枝子幼苗渗透调节物质的影响Fig.5 Effect of doubled CO2 on osmoregulatory substance of Lespedeza davurica seedling under drought stress
2.2.3干旱胁迫下CO2浓度倍增对达乌里胡枝子幼苗质膜透性和MDA含量的影响 图6a为CO2倍增和干旱胁迫对达乌里胡枝子幼苗相对电导率的影响。如图所示,随着自然干旱处理天数的延长,两种CO2浓度条件下达乌里胡枝子幼苗的相对电导率均上升,随着自然干旱处理天数的延长,两者间的差异逐渐变大,干旱胁迫至第10 d时,两种CO2浓度下的相对电导率开始出现显著差异(P<0.05),干旱胁迫至第16 d时,CO2倍增条件下的相对电导率较CK显著降低18.49%(P<0.05)。
图6b为CO2倍增和干旱胁迫对达乌里胡枝子幼苗MDA含量的影响。如图所示,干旱胁迫至第14 d时,两种CO2浓度条件下达乌里胡枝子幼苗的MDA含量均呈上升趋势,CO2倍增条件下的MDA均低于CK。干旱胁迫至第12 d时,两种CO2浓度条件下幼苗MDA含量差异显著(P<0.05),干旱胁迫至14 d时,CK条件下幼苗MDA含量有增加的趋势,而CO2倍增条件下MDA含量开始下降。干旱胁迫至第16 d时,CO2倍增条件的MDA含量比CK处理少了18.16%(P<0.05)。

图6 CO2倍增和干旱胁迫对达乌里胡枝子幼苗质膜透性和MDA含量的影响Fig.6 Effect of doubled CO2 on cytomembrane permeability and MDA contents of Lespedeza davurica seedling under drought stress
2.2.4干旱胁迫与CO2浓度倍增条件下达乌里胡枝子幼苗生理指标间的相关分析 由表1可知,幼苗在CK条件下的SOD与POD活性呈极显著正相关关系(P<0.01);SP含量与LRWC呈极显著正相关关系(P<0.01),与Pro含量呈显著负相关关系(P<0.05),与MDA含量和相对电导率呈极显著负相关关系(P<0.01);SS含量与Pro含量、相对电导率呈显著正相关关系(P<0.05),与LRWC呈显著负相关关系(P<0.05);Pro含量与LRWC呈极显著负相关关系(P<0.01),与MDA含量和相对电导率呈极显著正相关关系(P<0.01);LRWC与MDA含量、相对电导率呈极显著负相关关系(P<0.01);相对电导率与MDA含量呈极显著正相关关系(P<0.01)。CO2倍增处理下,幼苗SOD活性与POD活性呈极显著正相关关系(P<0.01),与MDA含量和相对电导率呈显著正相关关系(P<0.05);Pro含量与LRWC呈极显著负相关关系(P<0.01),与MDA含量和相对电导率呈极显著正相关关系(P<0.01);LRWC与MDA含量和相对电导率呈极显著负相关关系(P<0.01);MDA含量与相对电导率呈极显著正相关关系(P<0.01)。
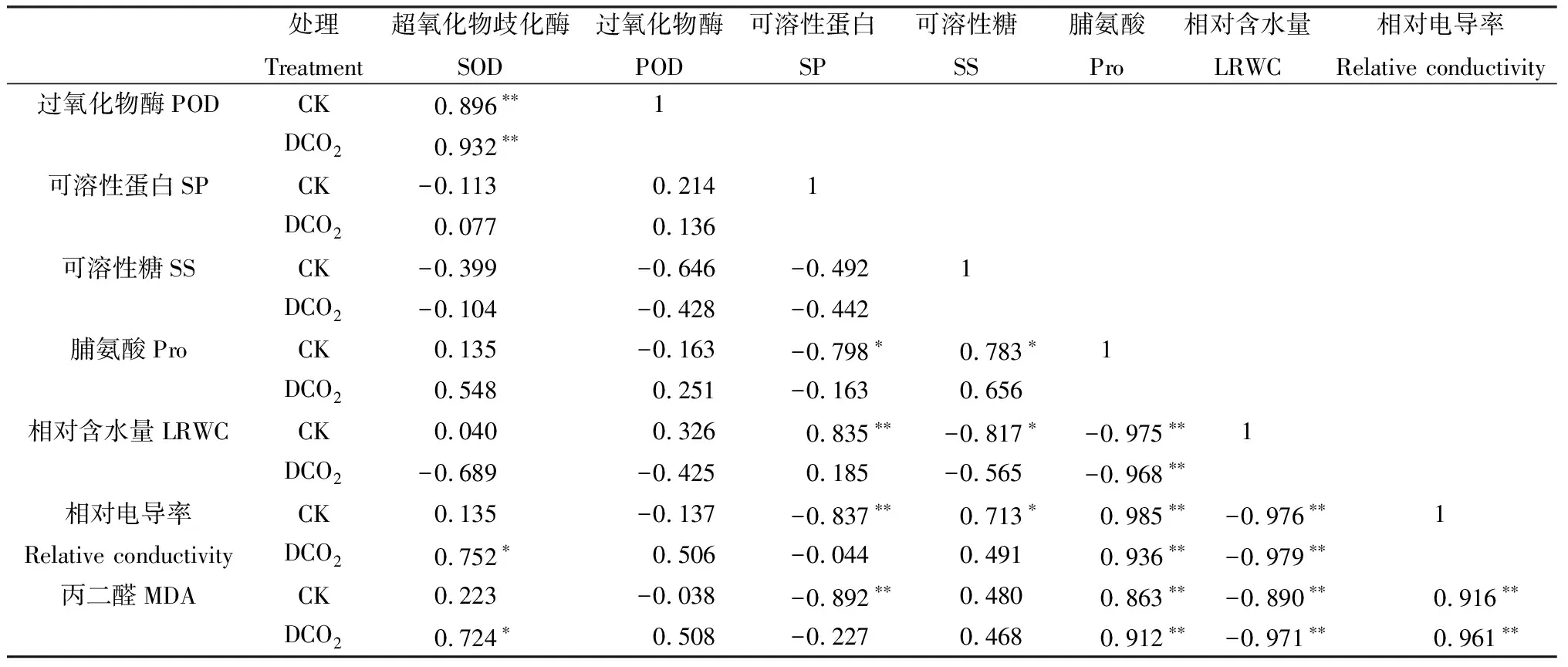
表1 对照和CO2倍增条件下达乌里胡枝子幼苗各生理指标间的相关分析Table 1 Correlation analysis of physiological indexes of Lespedeza davurica seedlings
3 讨论
3.1 干旱胁迫与CO2浓度倍增条件下达乌里胡枝子幼苗的生长特性
干旱胁迫会导致植物生长所需水分的吸收量不足,导致植株地上部分生长缓慢,为了维持植株正常生长,植株会促进地下根系的生长,以便获取更多水分供生命活动,而CO2浓度升高能够促进地上部和地下部生物量的增加[28]。本试验研究结果表明:短期的自然干旱会抑制达乌里胡枝子幼苗植株地上部分的生长,在干旱胁迫8~12 d时抑制作用缓解,在CO2浓度倍增条件下,自然干旱的幼苗株高相对生长速率高于CK处理,而且在干旱胁迫初期时会对达乌里胡枝子的幼苗地上部分起促进作用,说明在一定程度的干旱胁迫下高浓度的CO2会促进植株幼苗地上部分的生长;自然干旱会对幼苗地下部生长起促进作用,干旱程度加深时这种促进作用较明显,这是因为在干旱条件下CO2浓度升高能促进地上部营养物质合成并向根部运输,促进根系伸长以便于获取更深层的水分。杨敏慎等[29]研究表明气候变暖和CO2浓度升高对农作物地上和地下生物量及产量都具有促进作用。杨娟等[30]研究认为,干旱胁迫下玉米(Zeamays)苗期地下根系比地上部分响应更为敏感,均与本研究结果相似。
3.2 干旱胁迫与CO2浓度倍增条件下达乌里胡枝子幼苗的抗氧化特性
研究表明,干旱胁迫下植物体内发生膜脂过氧化,产生活性氧自由基后破环了细胞膜,导致细胞功能的紊乱,诸多植物为了应对干旱胁迫对本体造成的伤害,会促进相关抗氧化酶的合成,提高相关酶的活性,以缓解干旱胁迫给植株带来的伤害[31-33]。本研究表明:在一定时间内干旱胁迫,可促进幼苗中SOD和POD活性的升高,随着胁迫程度加深(10~16 d)幼苗的SOD和POD活性均受到抑制呈下降趋势;CO2浓度倍增情况下在一定干旱胁迫时间内(2~12 d)可以更好的促进SOD和POD酶活性的升高,这样可以减轻干旱胁迫对植株的伤害[34],但过度干旱会伤害植物抗氧化机制导致抗氧化酶活性降低[35]。相较于CK,CO2浓度倍增能在更长时间干旱胁迫提高达乌里胡枝子幼苗体内SOD和POD活性,说明高浓度CO2能在更大干旱程度上促进植物体内抗氧化酶活性的提高,较CK更有效的缓解干旱胁迫,较大程度提高植物的抗旱能力。李仪曼等[36]通过增加CO2浓度提高了干旱胁迫下黄瓜幼苗的根系抗氧化酶系统的防御能力,缓解了干旱胁迫对黄瓜幼苗的伤害,增强其耐旱能力保证幼苗生长,通过研究表明,CO2浓度倍增可以提高植物幼苗的抗旱性,有利于植物更好的应对未来全球气候变化所带来的干旱影响。
干旱胁迫诱发植物体内膜脂过氧化,促进膜磷酸脂脱酯化反应,加速膜结构和功能的破坏,植物体内所含渗透调节物质的多少可反映其所受干旱胁迫的程度[37]。本试验研究结果表明:在轻度干旱情况下能提高达乌里胡枝子幼苗叶片的SP和SS含量,当干旱程度加深时,SP含量降低,SS含量升高,CO2浓度升高对SP和SS含量没有明显的抑制和促进作用。干旱胁迫会促进达乌里胡枝子叶片Pro的积累,同时会降低叶片的LRWC,随着干旱胁迫程度的加深,效果越来越明显;CO2浓度升高可以提高叶片内的Pro含量和LRWC。刘遵春等[38]对金光杏梅幼苗叶片在干旱胁迫下的研究结果与本试验结果相似,说明SS和Pro是植物体内两种重要的渗透调节物质,当受到干旱胁迫时,植物体内会积累大量的SS和Pro,使植物保持一定的含水量和膨压,以维持细胞正常的功能。干旱胁迫能促进Pro含量的增加,在CO2浓度升高处理中度干旱和高度干旱下Pro含量呈显著性上升。高浓度CO2能促进植株进行光合作用,减少干旱胁迫下植株叶片的水分散失,在干旱环境中保持相对较高的LRWC,缓解干旱胁迫对植株的危害。
植物遭受干旱胁迫时MDA含量发生变化,发生膜脂过氧化进而破坏膜结构,导致细胞膜的选择透过性下降,MDA含量的高低可以反映植物所受逆境胁迫程度的大小。本研究表明:干旱程度较轻时(2~8 d)达乌里胡枝子幼苗相对电导率无明显变化,干旱程度较重时(10~16 d)幼苗叶片相对电导率升高,CO2浓度倍增条件下叶片相对电导率显著低于CK;在短期干旱时间内(2~10 d)MDA含量变化不明显,随着干旱胁迫时间的增加,叶片中MDA含量明显增加。干旱胁迫短时间内,达乌里胡枝子幼苗的相对电导率和MDA含量上升幅度较小,这是因为达乌里胡枝子幼苗本身具有一定耐旱性。当胁迫程度超过了达乌里胡枝子本身的耐旱阈值,会破坏细胞结构,导致幼苗的相对电导率和MDA含量明显上升。与CK相比CO2倍增处理降低了幼苗的相对电导率和MDA含量,是因为高浓度CO2能够缓解干旱条件下膜脂过氧化的发生,保护生物膜系统,降低MDA含量的累积与植物细胞的相对电导率。李清明等[39]研究说明CO2浓度升高可显著促进干旱胁迫下尤其是重度干旱情况下黄瓜幼苗叶片渗透物质的积累,与本研究结果一致,认为CO2浓度升高可在一定程度上减轻黄瓜幼苗干旱胁迫下活性氧积累造成的伤害,维持活性氧的平衡,致使质膜相对透性降低,MDA含量减少,对防止植物的氧化损伤具有一定的保护作用,忽雪琦等[40]对玉米幼苗根系的研究结果也与本试验结果相似。
4 结论
本试验探讨在干旱胁迫时CO2浓度倍增对‘晋农1号’达乌里胡枝子幼苗生长特性和抗氧化特性的影响,CO2浓度倍增缓解了干旱胁迫对达乌里胡枝子幼苗株高的抑制作用,促进其主根的生长。CO2浓度倍增提高了干旱胁迫下达乌里胡枝子幼苗的SOD和POD活性,提高了幼苗的抗氧化酶活性,增加了LRWC和Pro含量,抑制MDA合成,降低了相对电导率,延缓了膜脂过氧化的发生。综上所述,CO2浓度倍增减轻了干旱对植物细胞的伤害,提高了达乌里胡枝子幼苗对干旱的耐受性,同时也提高了达乌里胡枝子的生产性能。
